TCA 回路は単なるエネルギーの製造装置ではありません。様々な生体分子の前駆体を与えるという役割も担っています。このことは制御を見ていくことでよく理解できると思います。今回は TCA 回路の制御について解説していくと同時に、他の代謝経路との関係性について解説していこうと思います。
Contents
制御のポイントは三つ、TCA の入り口と二つの NADH 生成段階で調節される
TCA 回路の制御ポイントは三つです。まず一つ目はピルビン酸デヒドロゲナーゼで、この反応は TCA 回路に入る前段階です。さらにもう二つはイソクエン酸デヒドロゲナーゼと 2-オキソグルタル酸デヒドロゲナーゼの二段階です。イソクエン酸デヒドロゲナーゼはイソクエン酸から 2-オキソグルタル酸の変換段階を触媒し、2-オキソグルタル酸デヒドロゲナーゼは 2-オキソグルタル酸からスクシニル CoA の変換段階を触媒します。この二つの段階では NADH を産生する段階です。それでは各段階の制御についてみていきましょう。
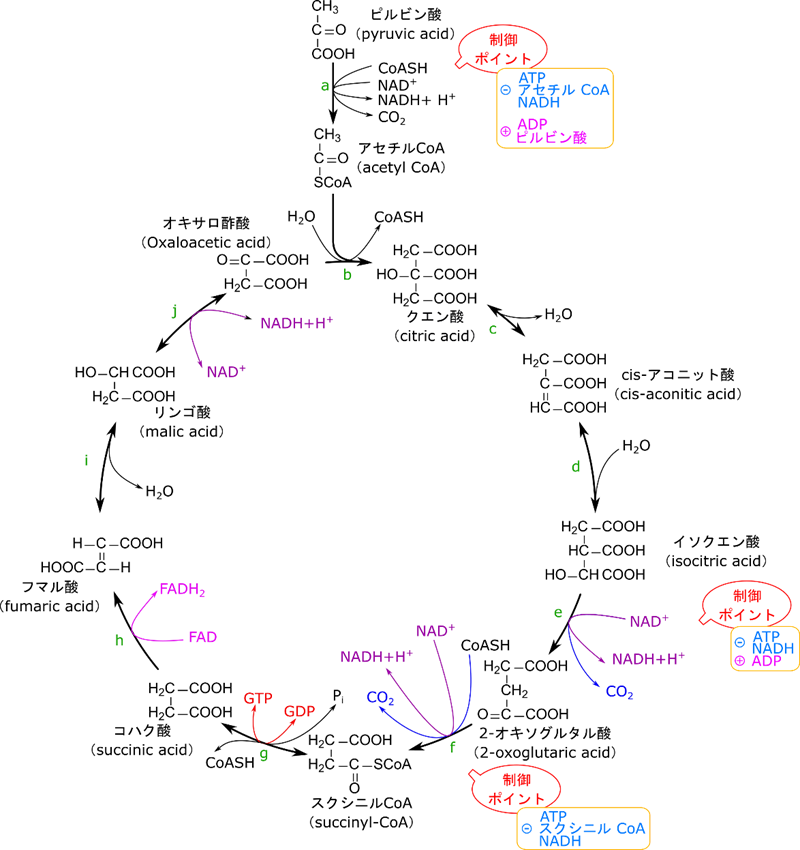
ピルビン酸デヒドロゲナーゼに対する制御はエネルギーの充足率に応じて行われる
ピルビン酸がアセチル CoA になるとグルコースに戻すことはできません。ですので、必要がない場合にはこの段階でストップをかけることはとても重要です。アセチル CoA から先には
- TCA 回路に入って炭酸に異化される
- 脂肪酸合成に入って貯蓄される
の二通りです(図3)。脂肪酸合成に向かうべき状況とはどういう状況でしょうか? 上記の2経路について考察すると
- TCA 回路はエネルギーを生み出す経路
- 脂肪酸合成はエネルギーの貯蔵をする経路
ですので、
- エネルギーが不足するときには TCA 回路を誘導するべき
- エネルギーが充足していると脂肪酸合成を誘導するべき
となります。つまりエネルギーが充足している場合は TCA 回路は抑制されるべきであるといえます。
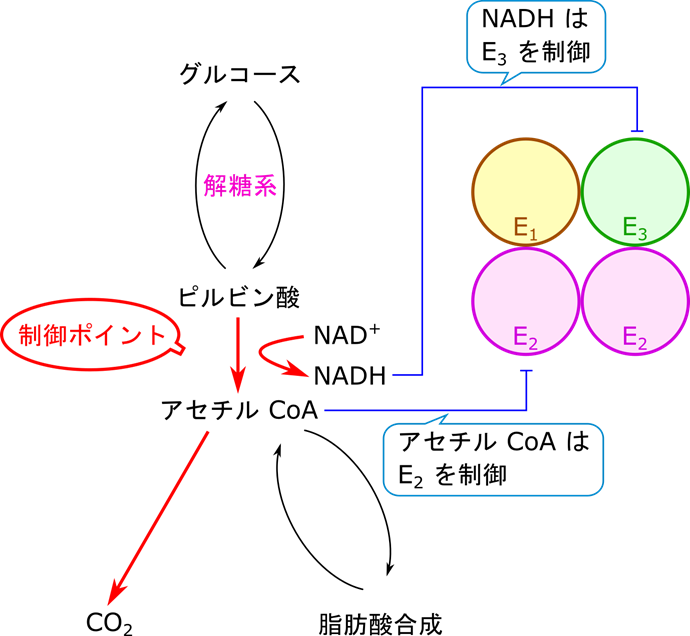
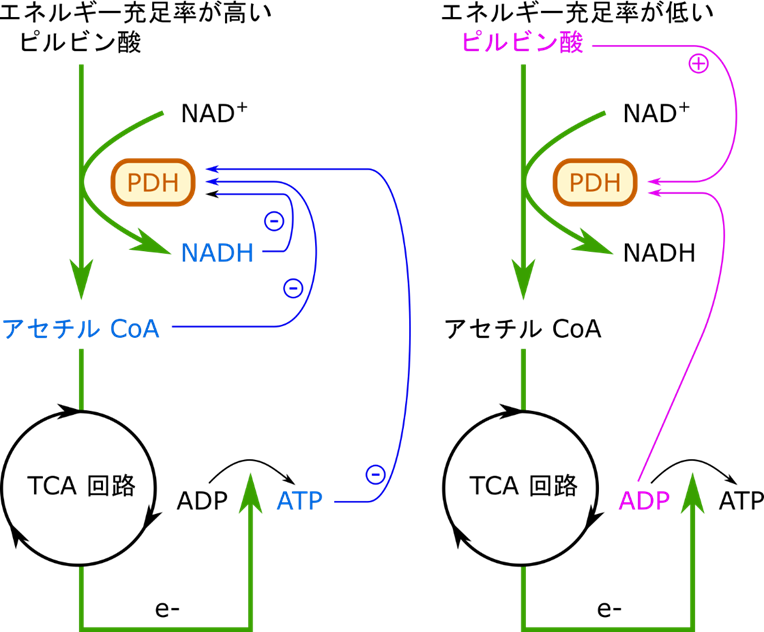
NADH は ATP に変換されるわけですので、NADH が高濃度になる場合はエネルギーが充足している状況と理解できます。NADH はピルビン酸デヒドロゲナーゼの E3 サブユニット(ピルビン酸デヒドロゲナーゼは複合体 E1 ~ 3 の三種類のタンパク質の複合体になっています)に直接結合して阻害します。では、アセチル CoA が高濃度の状況はどうでしょうか?この場合は少し事情が異なります。実はアセチル CoA の供給源はピルビン酸だけではありません。脂肪酸がエネルギーにされる際にもアセチル CoA が供給されます。つまり、アセチル CoA が豊富である状況というのは脂肪酸が分解されてアセチル CoA が供給されている状況を表しています。この場合は当然解糖系からアセチル CoA を供給する必要はありませんね。ですのでアセチル CoA は E2 サブユニットに直接結合してピルビン酸デヒドロゲナーゼの機能を抑制します。
一方でピルビン酸デヒドロゲナーゼの制御はキナーゼとホスファターゼを介しても行われます。キナーゼはリン酸を付ける酵素ですね。一方でホスファターゼはリン酸を取り去る作用を持つ酵素です(図2)。ピルビン酸デヒドロゲナーゼ複合体はリン酸化を受けるとその活性がが抑制され、不活性型になります。一方でホスファターゼが作用するとリン酸が除去されてピルビン酸デヒドロゲナーゼ複合体は活性化されます(図2)。ではどういった因子がホスファターゼやキナーゼの活性を誘導するのでしょうか?ATP/ADP 比、アセチル CoA/CoA 比、NADH/NAD 比が上昇する(高ATP、高アセチル CoA、高 NADH 条件ということになります)と、キナーゼ(ピルビン酸デヒドロゲナーゼキナーゼ)が活性化されて、ピルビン酸デヒドロゲナーゼがリン酸化されて不活性化されます。これは高エネルギーや脂肪酸分解が進行している状況であることは上述の通りです。このような場合は TCA 回路を動かす必要はなかったり、ピルビン酸からアセチル CoA を供給する必要がないためピルビン酸デヒドロゲナーゼの活性が抑制されますね。一方で、ADP やピルビン酸濃度が高まるとキナーゼの作用を阻害します。一方でこのような状況とはエネルギーが使われている状況であり、筋収縮が行われている状況です。筋収縮は Ca2+ が流入することによって起こりますが、このシグナルは同時にホスファターゼも活性化されます。これらの連携によってピルビン酸デヒドロゲナーゼは活性化されます。さて、当然ではありますがこのような状況はエネルギーが必要とされている状況なので TCA 回路を動かすべき状況ですよね。
ではいったんまとめます。図4を見てください。NADH や ATP が高濃度になると、エネルギーが充足しているので、ピルビン酸デヒドロゲナーゼの活性は抑制されます。また、アセチル CoA が高濃度になると、脂肪酸が分解されてアセチル CoA が供給されていることを意味するので、ピルビン酸デヒドロゲナーゼの活性は抑制されます。一方でピルビン酸や ADP が高濃度になると、エネルギーを要求していることになるので、ピルビン酸デヒドロゲナーゼの活性は増大させられます(念のためピルビン酸が高濃度になる場合について説明します。ピルビン酸は解糖系の最終産物ですよね。解糖系はエネルギーを要求される場合に機能する代謝経路です。ですので、エネルギーが必要とされる状況(運動期)にはピルビン酸が活発に産生されることになります)。

二つの NADH 生成段階の制御はエネルギーの充足度合いにより調整される
イソクエン酸回路の制御では ATP や NADH がイソクエン酸デヒドロゲナーゼの作用を阻害します。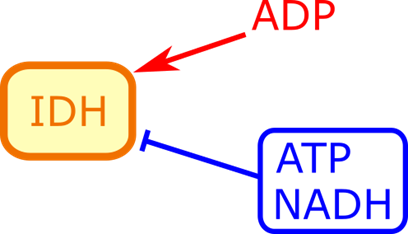
逆に ADP はイソクエン酸デヒドロゲナーゼの基質との親和性を高めて機能を促進します。イソクエン酸デヒドロゲナーゼの活性が抑制されると基質の消費が滞るのでイソクエン酸が蓄積しますね。するとクエン酸→イソクエン酸の反応は可逆的ですので、蓄積したイソクエン酸はクエン酸まで戻ってしまいます。ところがクエン酸から以前には戻れませんのでクエン酸が蓄積しますね。すると蓄積したクエン酸は解糖系を抑制することで解糖系からピルビン酸が流入することを停止します(図5)。
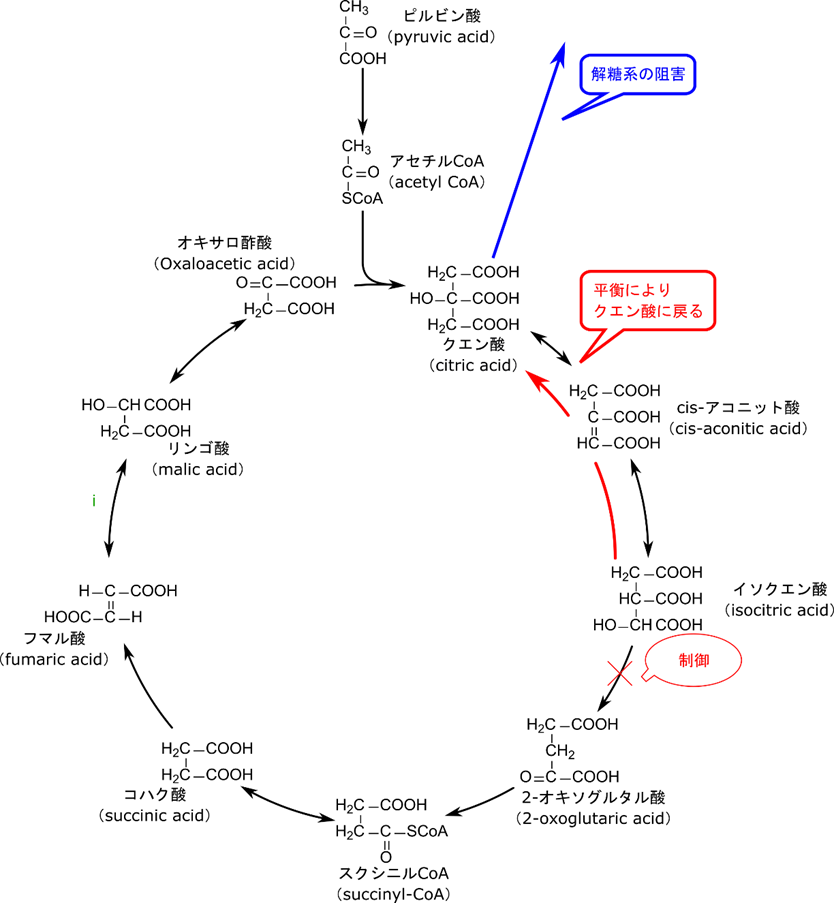
2-オキソグルタル酸デヒドロゲナーゼの活性は ATP、NADH、スクシニル CoA の作用によって阻害されます。![]()
この酵素はピルビン酸デヒドロゲナーゼと非常によく似た酵素です。実際反応も
ピルビン酸 + NAD+ + CoA → アセチル CoA + CO2 + NADH
2-オキソグルタル酸 + NAD+ + CoA → スクシニル CoA + CO2 + NADH
とよく似ていますね。ですので制御も似てきます。つまり、ピルビン酸デヒドロゲナーゼは ATP と NADH、反応生成物(アセチル CoA)によって抑制されますが、2-オキソグルタル酸デヒドロゲナーゼでも同様に ATP と NADH、反応生成物(スクシニル CoA)によって阻害されます。これまでも制御には生理的な意味がありました。当然 2-オキソグルタル酸デヒドロゲナーゼの制御にも生理的な意味があります。2-オキソグルタル酸デヒドロゲナーゼが抑制されると 2-オキソグルタル酸が蓄積されます。ところがイソクエン酸から 2-オキソグルタル酸の反応は不可逆反応ですのでいくら 2-オキソグルタル酸が蓄積しても戻っていきません。すると 2-オキソグルタル酸が蓄積する一方になってしまいます。こうして蓄積した 2-オキソグルタル酸はアミノ酸合成やアミノ酸合成を介したプリン塩基の合成に使用されます(図6)。
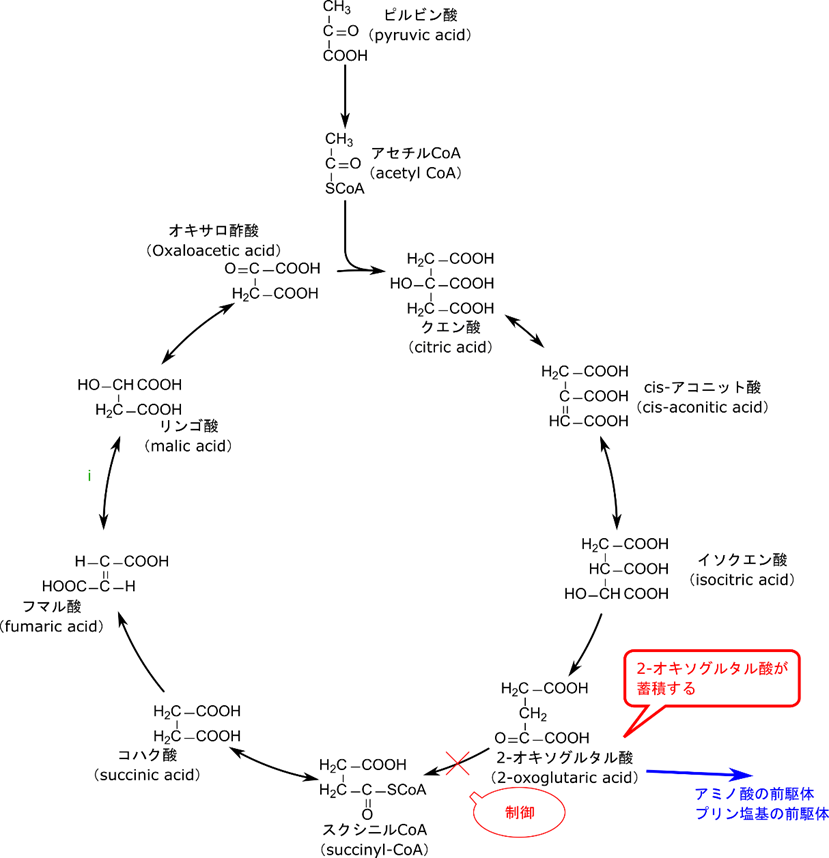
TCA 回路は他の代謝経路の前駆体を供給する
TCA 回路はエネルギー供給のための代謝経路というイメージが強いですが、他の生体分子の前駆体を供給する役割も担っています(図7)。順番に見ていきましょう。
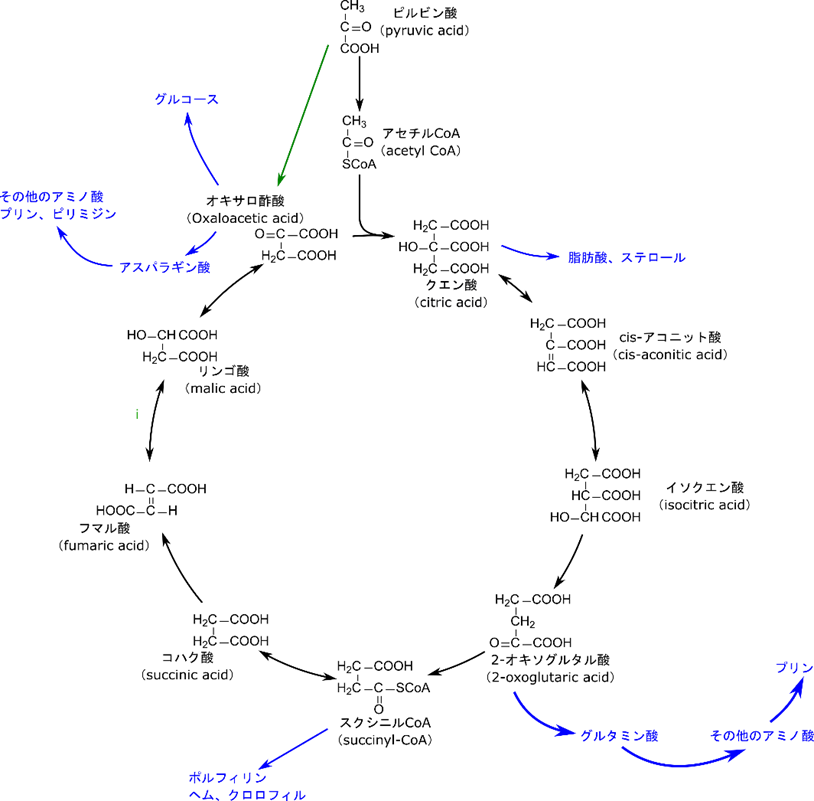
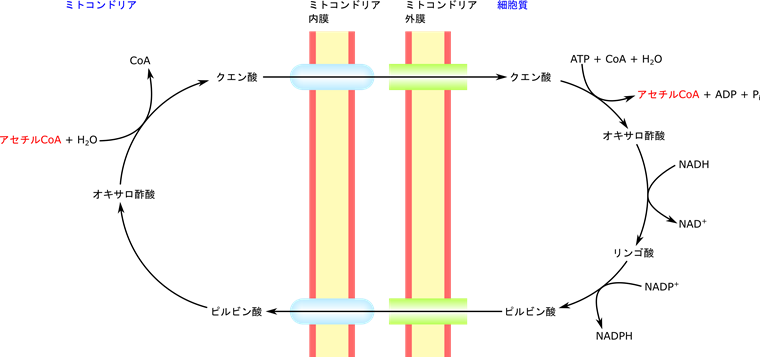
まず、クエン酸は脂肪酸やステロールの前駆体となります。脂肪酸やステロールの合成にはアセチル CoA が必要となります。このアセチル CoA は TCA 回路の前段階、ピルビン酸デヒドロゲナーゼによって合成されますね。この反応も TCA 回路も、ミトコンドリアのマトリックス内で行われます。一方で、脂質合成は細胞質で行われます。このためアセチル CoA をミトコンドリア外に運ばないといけません。ところがアセチル CoA はミトコンドリアの内膜を通過できません。これではアセチル CoA を利用することができません。この時に運搬役を担うのがクエン酸です。クエン酸がミトコンドリア内膜を通過して細胞質に運搬されます。するとクエン酸が分解されると同時に CoA と縮合されてアセチル CoA が合成されます。このようにしてアセチル CoA が細胞質に運搬されました(図8)。このアセチル CoA が脂肪酸やステロール合成に使用されます。以上がクエン酸が脂肪酸、ステロールの前駆体として使用されるということの意味です。
次は制御の項目でも記載しましたが、2-オキソグルタミン酸です。アミノ基の取り込みはグルタミン酸がアンモニアを受け取り、グルタミンに変換されるところから始まります。グルタミンは 2-オキソグルタル酸にアミノ基を渡してグルタミン酸に変換されます。一方で 2-オキソグルタル酸もグルタミン酸に変換されます。さらに合成されたグルタミン酸はアミノ基を様々な α-ケト酸に渡すと、種々のアミノ酸を合成します。するとグルタミン酸は 2-オキソグルタル酸に変換されます。このようにして 2-オキソグルタル酸は種々のアミノ基の取り込みに関与して種々のアミノ酸の前駆体となります(図9)。

プリンの合成は何段階もの反応を経て合成されます。合成経路については複雑なのでここでは記載を避けますが、プリンの各原子の由来を以下に示します。グルタミンはグルタミン酸から合成されますが、グルタミンのアミノ基がプリンの窒素を供与します(図10)。
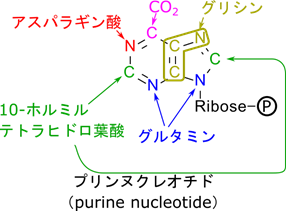
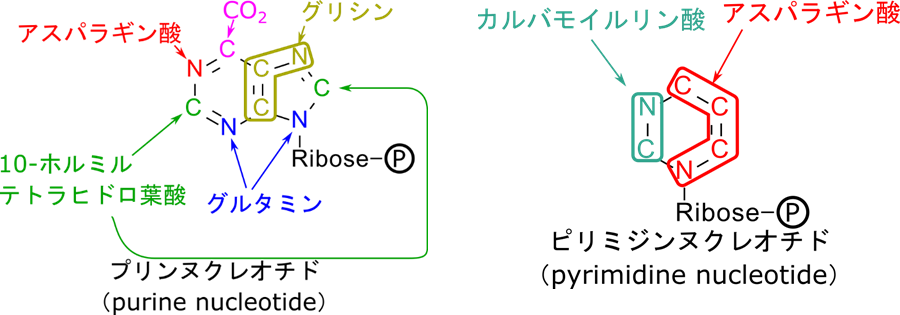
2-オキソグルタル酸と同様にオキサロ酢酸もアミノ酸、プリン、ピリミジンの前駆体を供与します。まずはアスパラギン酸の合成を記載します(図11)。この反応は図9の α-ケト酸のところにオキサロ酢酸を当てはめて考えてみてもらえたらわかると思います。ところで、グルタミン酸、グルタミン、アスパラギン酸というのは他のアミノ酸を合成する出発物質ともなります。この意味で、2-オキソグルタル酸とオキサロ酢酸はアミノ酸全体の前駆体とも言えます。

さらに、プリン、ピリミジンを構成する原子の由来を示すと以下のようになります(図12)。このようにオキサロ酢酸はアスパラギン酸を介してプリン、ピリミジンの原子を供与します。
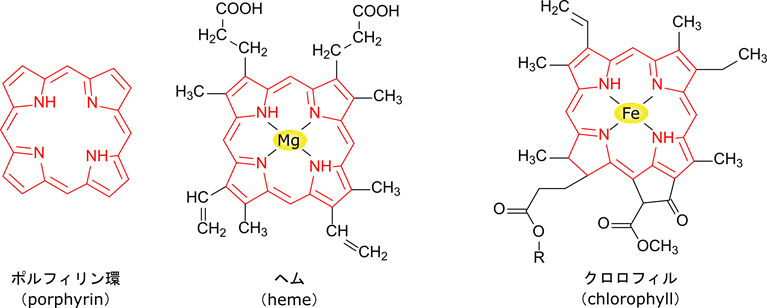
次に、スクシニル CoA です。ポルフィリン環はヘムやクロロフィルのような金属錯体を形成するような分子に含まれる構造です(図13)。ポルフィリン環の生合成はここでは記載しませんが、スクシニル CoA とグリシンから何段階かの反応経て合成されます。
ところで、これまで解説したように TCA 回路の中間産物は他の代謝経路に前駆体として供給されていきます。すると、エネルギーを供給しなければならないときに TCA 回路を動かすことができなくなってしまう可能性があります。というのも、回路を動かすためにはオキサロ酢酸がアセチル CoA を受け取らないといけないのですが、このオキサロ酢酸が大量に使われていたとしたらどうでしょうか? TCA 回路がうまく機能してくれなさそうですよね。このような場合にはピルビン酸が炭酸と縮合してオキサロ酢酸に変換されて供給されます。
ピルビン酸 + CO2 + ATP + H2O → オキサロ酢酸 + ADP + Pi + 2H+
このような代謝の中間産物を補充するような反応をアナプレロティック反応(anaplerotic reaction;ギリシャ語で “満たす” の意味です)といいます。こうすることにより TCA 回路の動きが鈍ってしまうことを防いでいます。
今回は、TCA 回路の制御と他の代謝経路との関係についてみてみました。このように見てみると TCA 回路はただ単なるエネルギー産生経路というだけでなく、様々な分子の出発物質を与えるとても基本的な合成経路でもあります。その意味で他の経路を考える上でもとても大切な代謝経路ですのでぜひしっかりと勉強してみてください。
参考文献
- 島原健三 (1991). 概説 生物化学. 三共出版. pp. 151-162
- Jeremy M. Berg, John L. Tymoczko, Gregory J. Gatto Jr., Lubert Stryer著、入村達郎、岡山博人、清水孝雄、中野徹訳 (2018). ストライヤー生化学 第8版. 東京化学同人. pp. 416-442
- John McMurry著、伊東椒、児玉三明訳 (2000). マクマリー有機化学 第4版. 東京化学同人. pp. 365-367, 546-551
- Paula Y. Bruice 著、大船泰史、香月勗、西郷和彦、富岡清訳 (2016). ブルース有機化学概説 第3版. 化学同人. pp. 678-729
関連記事
糖鎖の基本について
糖鎖の化学反応について
オリゴ糖・多糖について
解糖系について
- 解糖系(glycolysis)によって糖からエネルギーを取り出しATP を生成する
- 解糖系の各反応をエネルギー的観点から読み解く
- 解糖系で利用される糖はグルコースに限らない
- 解糖系の制御は三つの律速酵素に対する調節で行われる
TCA サイクルについて