解糖系は糖からエネルギーを取り出す最も基本的な代謝経路の一つです。原核生物から私たちヒトに至るまで多くの生物が持っている代謝経路で、もっとも原始的な代謝経路でもあります。解糖系についてはご存じの方も多いかと思いますが、糖を分解するだけでなぜエネルギーが取り出せるのか疑問に思ったことはないですか?実は系の中にエネルギーを取り出すための工夫が隠されています。ここでは解糖系の中でも特に注意すべき反応や重要な反応について解説していこうと思います。
解糖系の全容は?反応の概略についてみていきましょう
以下に解糖系の概略を記載します。複雑な系なのでまずは全体を把握してから各論を確認していきましょう。
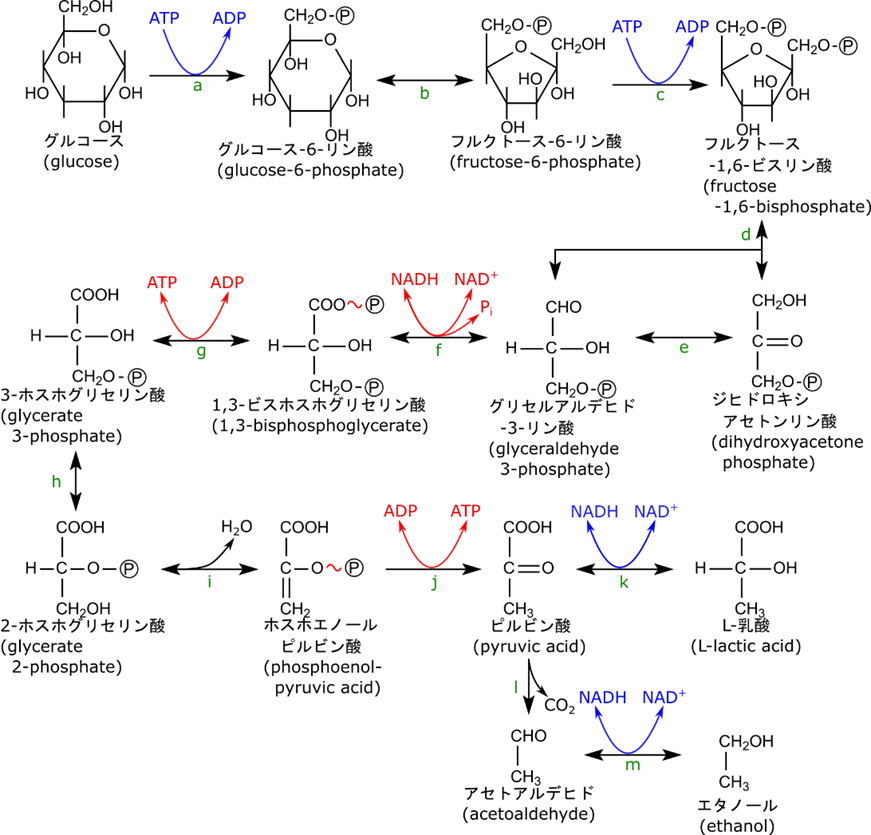
各反応を触媒する酵素名
a. グルコキナーゼ(ヘキソキナーゼ) b. グルコースリン酸イソメラーゼ c. ホスホフルクトキナーゼ d. フルクトースビスリン酸アルドラーゼ e. トリオースリン酸イソメラーゼ
f. グリセルアルデヒドリン酸デヒドロゲナーゼ g. ホスホグリセリン酸キナーゼ
h. ホスホグリセリン酸ムターゼ i. エノラーゼ j. ピルビン酸キナーゼ k. 乳酸デヒドロゲナーゼ
l. ピルビン酸デカルボキシラーゼ m. アルコールデヒドロゲナーゼ
グルコースからエネルギーを取り出す手順についてみていきましょう。解糖系は前半と後半に分けて考えることができます。
前半は、グリセルアルデヒド-3-リン酸までで、グルコースを半分に切断してグリセルアルデヒド-3-リン酸二分子を得ています。このようにすることで、後半のエネルギー取り出し過程を2分子分に増やすことができます。エネルギー取り出し過程ではグリセルアルデヒド-3-リン酸一分子で 2 ATP が産生されますが、グリセルアルデヒド-3-リン酸はグルコース一分子から二分子産生されるので、結局グルコース一分子から ATP は4分子合成されることになります。前半の過程で ATP が二分子消費されますが、全体として4分子の ATP が合成されるわけなので、全体の収支としては 2ATP が合成されたことになり、プラスになります。
後半では、1,3-ビスホスホグリセリン酸という高エネルギー分子、次いでホスホエノールピルビン酸という高エネルギー分子を産生することでエネルギーの取り出しを行っています。これらの分子はリン酸基との間にエネルギーを蓄えており、脱リン酸化をすることでエネルギーを取り出すことが可能です。したがって、グリセルアルデヒド-3-リン酸から 1,3-ビスホスホグリセリン酸の過程、3-ホスホグリセリン酸からホスホエノールピルビン酸の過程はそれぞれ 1,3-ビスホスホグリセリン酸やホスホエノールピルビン酸を得るための過程と考えてください。
酵素名の命名法について
ここで酵素の名前の付け方について簡単に解説しておきましょう。表1に解糖系の各反応を触媒する酵素名と反応の種類をまとめています。以下の解説と見比べながら読んでいただけると理解しやすいかと思います。酵素の名前は前半に基質、後半に酵素の種類を記載します。酵素の種類は反応名に続けて語尾を -ase にします。例えば a の反応を媒介するヘキソキナーゼです。キナーゼというのはリン酸基を転位する反応を媒介する酵素です。前半は基質でしたね。この場合の基質はグルコースです。ヘキソースですね。なので “ヘキソ” が基質を意味します。ですのでヘキソキナーゼとなるわけです。他の酵素名も同様にしてみていただければ、酵素名が理解しやすいと思います。さて、注意すべきはホスホグリセリン酸キナーゼです。この酵素は反応 g を媒介していますが、反応産物は 3-ホスホグリセリン酸です。基質ではなく産物が名前になっていますね。これはキナーゼがリン酸基を ATP から別の分子に転位させる酵素なので、反応 g を逆側から見て名前を付けているからです。つまり、
を逆から見て
ととらえて命名しているのです。こう見ると ATP からリン酸を転位する反応と見ることができますね。すると基質は 3-ホスホグリセリン酸、酵素はキナーゼということでホスホグリセリン酸キナーゼという命名になります。酵素の命名ではその酵素名が本来意味している反応と逆になってしまうことがあります。このような場合は逆の反応ととらえて命名される場合がありますので注意してください。同様の命名が反応 j を触媒するピルビン酸キナーゼについても使われています。
解糖系反応の各論
この項目では解糖系の反応の内でもポイントになる反応についてみていこうと思います。前項目の概要を頭に入れたうえで読んでいただけるとより分かりやすいと思いますので、上の記事と見比べながら読んでみてください。
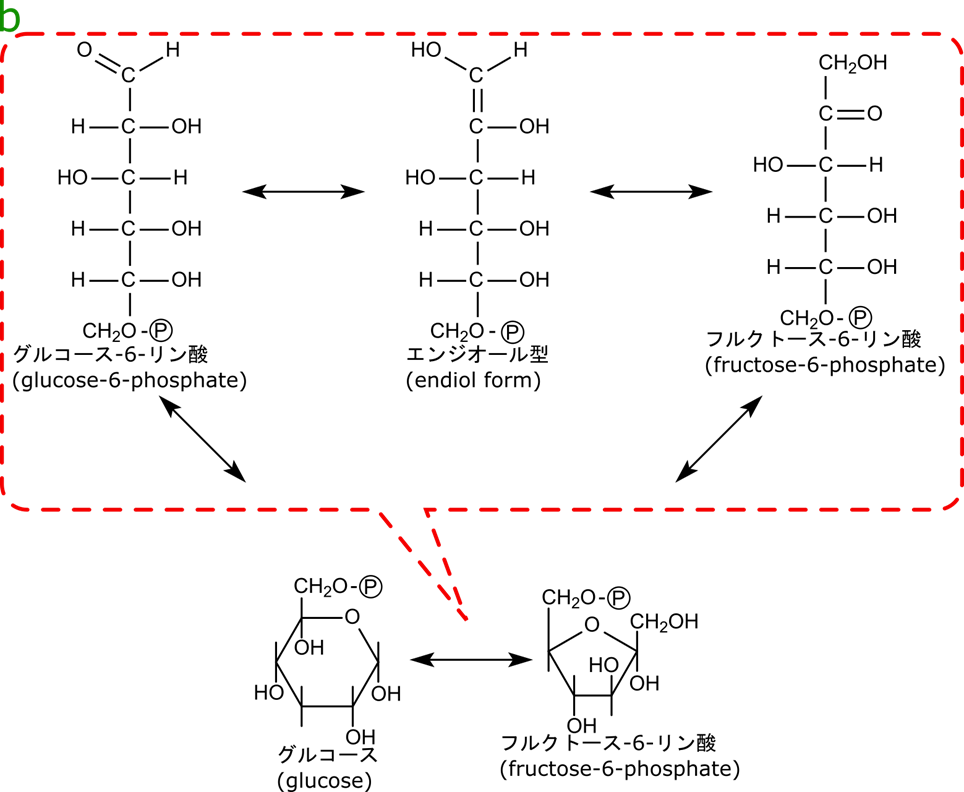
グルコース-6-リン酸からフルクトース-6-リン酸の異性化はエンジオールを介して行われる
このステップは、六炭糖を半分の位置で開裂するために必要なステップです。次の項目で詳述しますが、六炭糖を開裂する位置はカルボニル基(C=O)の位置に左右されます。グルコースからフルクトースへの異性化はカルボニル基の位置を調整する過程と理解してください。
グルコース-6-リン酸からフルクトース-6-リン酸への変換は、まず鎖状グルコースに開環します。次にエンジオール(C=C の両側に水酸基がひとつずつ着いた構造)を介してフルクトースに変換されます。その後形成されたフルクトース-6-リン酸は環状構造をとります。こうしてグルコース-6-リン酸からフルクトース-6-リン酸が生成されます。ちなみにこの反応はケト-エノール互変異(keto–enol tautomerism)と呼ばれる反応です(この反応はこちらの記事でも紹介していますので読んでみてください)。
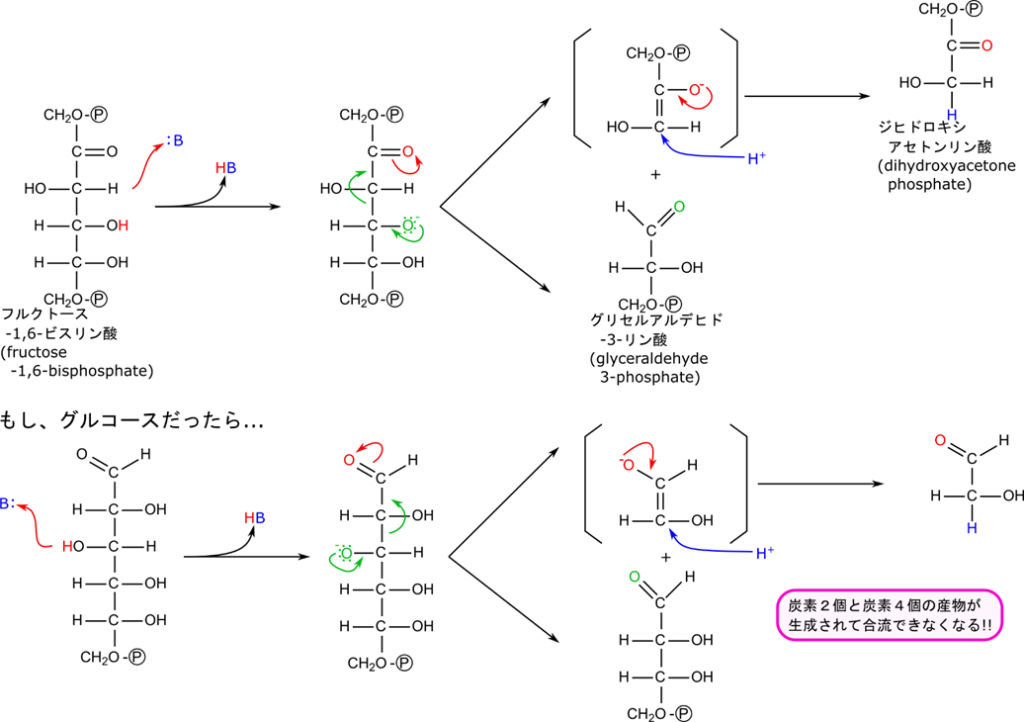
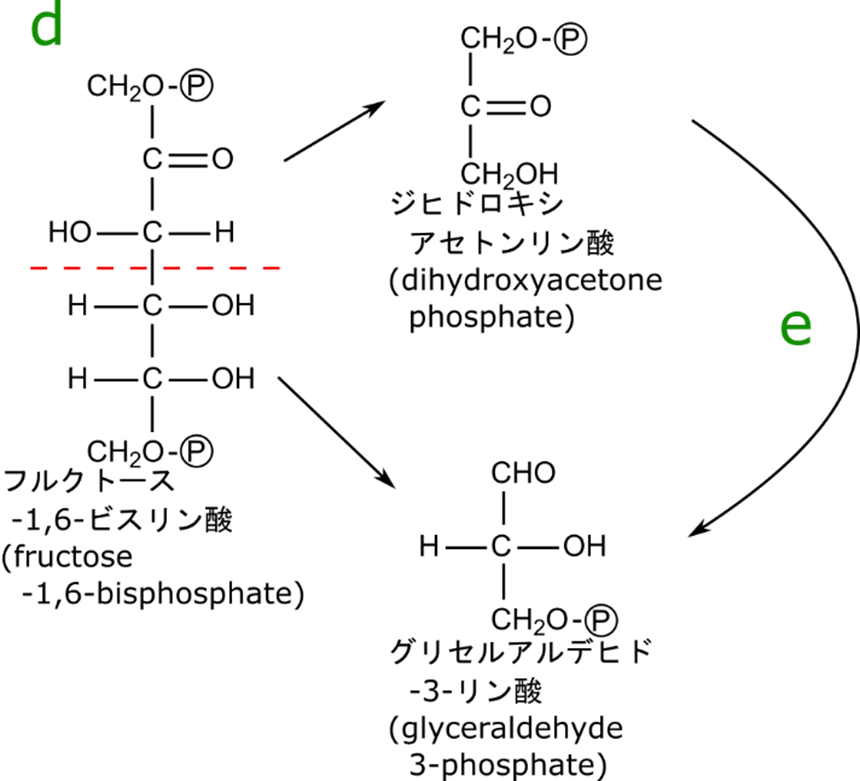
フルクトース-1,6-リン酸は開裂して3炭糖になる
フルクトース-1,6-ビスリン酸の開裂はジヒドロキシアセトンリン酸とグリセルアルデヒド3リン酸を生成します。産生されたジヒドロキシアセトンリン酸は異性化してグリセルアルデヒド3リン酸に変換されます(図4)。
まずはフルクトース-1,6-ビスリン酸の開裂を見ていきましょう。この反応はアルドール反応の逆反応になっています(アルドール反応については後述の「アルドール反応とは?」で紹介しています)。なお、アルドール反応を触媒する酵素なのでアルドラーゼと命名されています。
この反応は、カルボニル基の隣の炭素とその隣の炭素の間で開裂する反応です。まず、4位の水酸基からプロトンが塩基によって引き抜かれ、図3に示すように電子が遷移してグリセルアルデヒド-3-リン酸とエノラートアニオンが生成されます。エノラートアニオンにプロトンが付加してジヒドロキシアセトンリン酸が生成されます。
フルクトースへの変換が必要であることを検証するために、この反応をグルコースに直接作用させたと仮定してみましょう。図3の下側のような反応となり、炭素数が2個と炭素数が4個のアルデヒドが産生されてしまいます。このように、フルクトースへの異性化は六炭糖をちょうど半分の位置で開裂させるために必要な過程であることが確認できましたね。
さて、こうして生成されたジヒドロキシアセトンリン酸とグリセルアルデヒド3リン酸からリン酸をとった構造ジヒドロキシアセトンとグリセルアルデヒドはそれぞれ炭素数が3のケトースとアルドースです。ですので図1の e の反応はケトースからアルドースへの異性化であり、本質的に図1の b(アルドースからケトースへの異性化)の逆の反応とみなせます。つまり
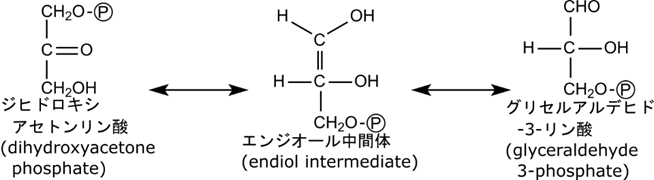
という経路を介してグリセルアルデヒド3リン酸に変換されるというわけです(当然この反応もケト-エノール互変異ですね)。ここで二種類の化合物がグリセルアルデヒド3リン酸で合流して共通の経路を利用できるようになります。
アルドール反応とは?
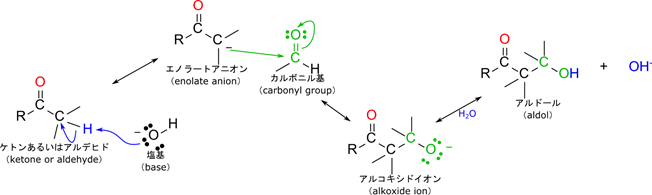
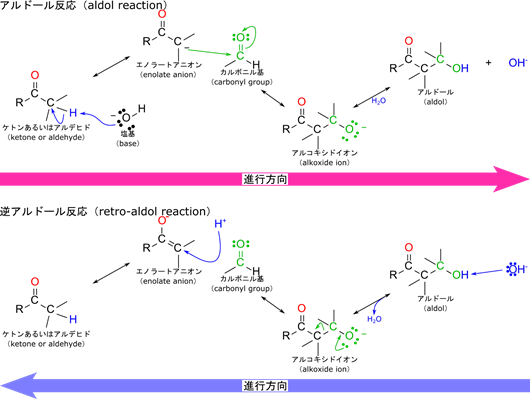
「フルクトース-1,6-リン酸は開裂して3炭糖になる」の説明の中に出てきたアルドール反応について概説します。必須というわけではないので難しいと感じる方は読み飛ばしてもらっても構いません。興味のある方は読んでみてください。
アルドール反応とはα 水素を持つカルボニル化合物をもう一つのカルボニル化合物と反応させるとこれら二つの化合物が連結されて β-ヒドロキシアルデヒドが産生される反応です。産生された物質は水酸基とアルデヒド基を併せ持つためアルドール(アルデヒド+アルコール)と呼ばれ、アルドールを生成することからアルドール反応(aldol reaction)と呼ばれます。反応メカニズムは上の図の通りです。まず、塩基が α 水素原子を引き抜いてエノラートアニオンが生成されます。このアニオンがカルボニル基の(δ+性を持つ)炭素に付加して結合を形成します。するとアルコキシドイオン中間体が生成されます。ここに水が付加するとアルドールが生成されます。
フルクトース-1,6-リン酸の開裂反応ではこの逆反応が発生します。このことをアルドール反応の逆の反応についてみていきながら確認してみましょう。塩基がプロトンを引き抜いてアルコキシドイオンが生成します。その後上の図のように電子が遷移してエノラートアニオンを生成しますが、この段階でカルボニルも生成します。プロトンが付加してケトン又はアルデヒドが生成します。フルクトース-1,6-リン酸の開裂の場合には、産生されるカルボニル化合物がグリセルアルデヒド-3-リン酸に相当し、エノラートアニオンはジヒドロキシアセトンリン酸に相当します。
グリセルアルデヒド3リン酸の酸化とリン酸化によって高エネルギー分子、1,3-ビスホスホグリセリン酸とNADH が産生される
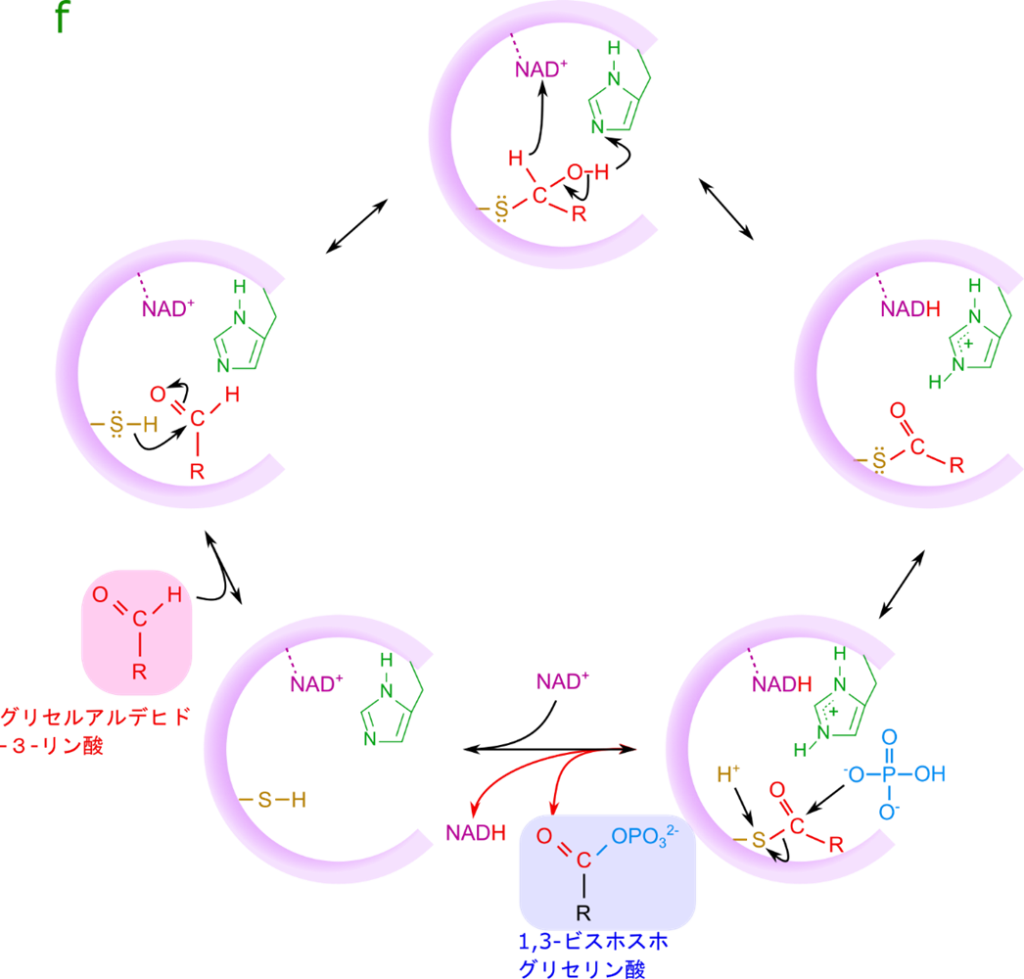
この反応はグリセルアルデヒド-3-リン酸を NAD+(ニコチンアミドアデニンジヌクレオチド;nicotinamideadenine dinucleotide)が酸化して(プロトンを奪って)、NADH と 1,3-ビスホスホグリセリン酸を産生する反応です(図5)。産生された NADH は電子伝達系によって ATP の産生に使用されます。その結果 NADH 1 分子当たり 3 分子の ATP が生成されます。ですので、この過程でも電子伝達系を介してエネルギーが産生されます。一方で産生された 1,3-ビスホスホグリセリン酸は高エネルギー分子です。なぜ高エネルギーなのかは次の項目で詳述します。さて、この反応がどのように触媒されるのかについてみていきましょう。
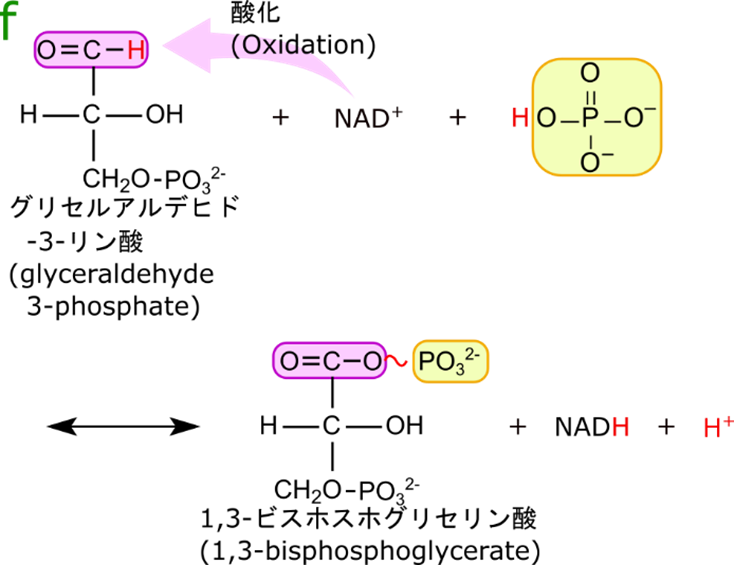
図6にグリセルアルデヒド3リン酸デヒドロゲナーゼの作用機序を記載しています。この酵素には NAD+ が結合しており、表面にチオール基(-SH 基)とヒスチジンが露出しています。ここにグリセルアルデヒド3リン酸が供給されるとチオール基の硫黄がアルデヒド基の炭素原子を求核的に攻撃して結合します。すると、プロトンが NAD+ に引き抜かれます。この過程では水酸基のプロトンがヒスチジンによって引き抜かれ、電子が遷移して水酸基(-OH)がカルボニル(-C=O)に変換されます。その結果、プロトンは押し出されて NAD+ による酸化が促進されます。チオエステル(硫黄を酸素に置き換えるとエステルですね。エステルの酸素の代わりに硫黄が利用されているのでチオ(硫黄)を付けてチオエステルといいます)が形成されます。ここにリン酸が付加されます。リン酸の酸素がチオエステルの炭素を求核的に攻撃して結合を形成しますが、これと同時に硫黄が外れてチオールに戻ります。この結果 1,3-ビスホスホグリセリン酸が生成されると同時に放出されます。また、NADH も放出されて、NAD+ と交換されます。さて、こうなると、酵素の状態は初期の状態に戻ります。ですので酵素は変化することなく、グリセルアルデヒド3リン酸から 1,3-ビスホスホグリセリン酸への変換が行われます。
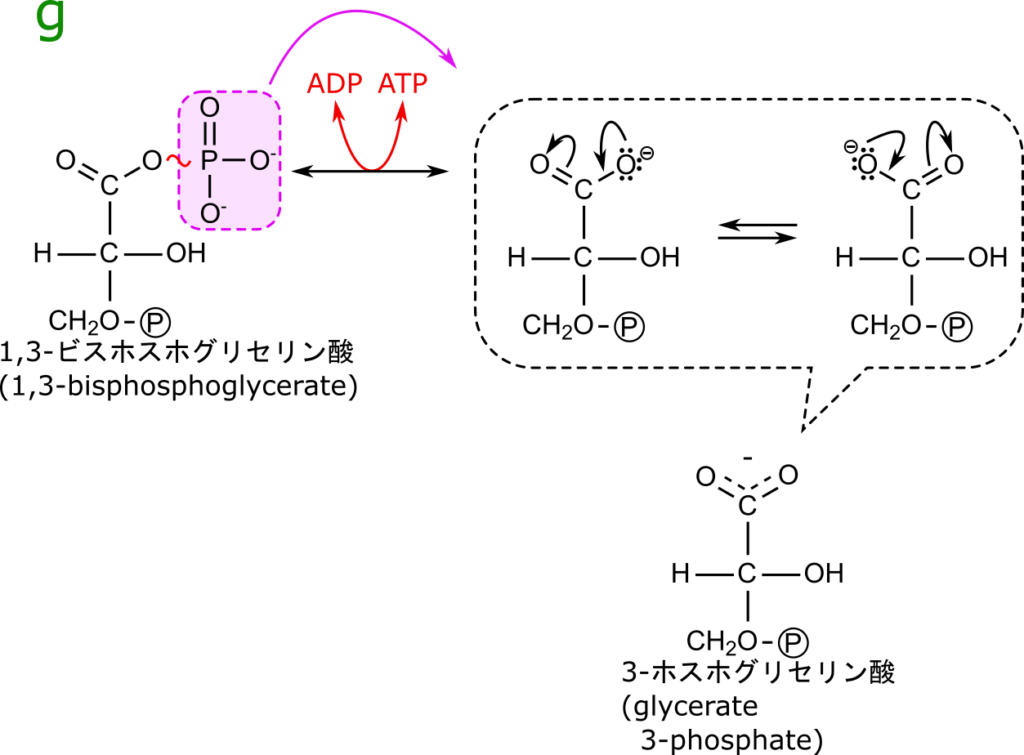
1,3-ビスホスホグリセリン酸は1位のリン酸基との結合が高エネルギー結合となっているため、リン酸を ADP に渡して ATP に変換することができます。ADP から ATP に変換する反応は吸エルゴン反応(エネルギーを要する反応)ですね。この時に必要となるエネルギーは1位のリン酸基を除去した際に得られたエネルギーを使用します。
ところで、1,3-ビスホスホグリセリン酸はなぜ高エネルギー分子なのでしょうか?特にリン酸基の結合が高エネルギー結合というのはどういうことなのでしょうか?図7を見てください。左側には 1,3-ビスホスホグリセリン酸の構造を、右側にはリン酸基をとった後の 3-ホスホグリセリン酸の構造を示しています。1,3-ビスホスホグリセリン酸ではエステルになっていますが、リン酸基が除去されて 3-ホスホグリセリン酸になるとカルボキシル基に変換されています。エステル結合では共鳴構造をとることはできませんが、カルボキシル基では共鳴構造をとることが可能です。このため 3-ホスホグリセリン酸の方が安定化します。この化学ポテンシャルの差分があるため 1,3-ビスホスホグリセリン酸からリン酸を除去するとエネルギーが取り出すことができるというわけです。
共鳴安定化することの意味とは?
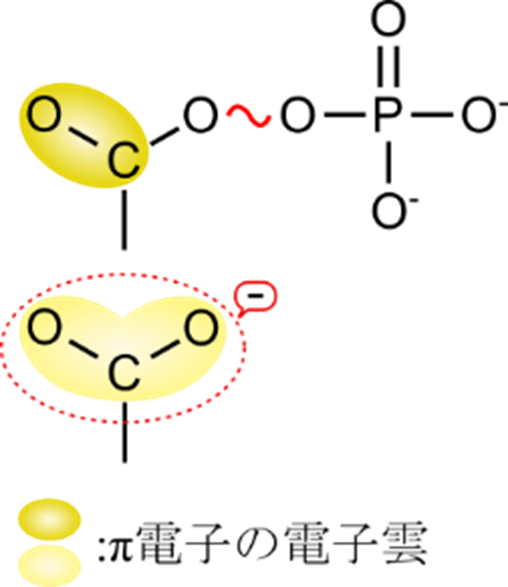
共鳴構造をとると安定化するという議論はよく見ると思います。しかし、なぜ共鳴構造をとると安定化するのでしょうか?ここからの記載はイメージとして理解していただければいいかと思いますので、そのつもりで読んでください。説明が求められた場合は「共鳴構造をとるため安定化する」という説明で十分です。
エステル結合をとっている場合、二重結合の π 電子は炭素-酸素間の結合以外の場所に存在することはできないため、狭い場所に押し込められています。電子雲でとらえると、C-O 間の電子密度が高くなります。一方で、カルボキシル基では、図7のように共鳴構造をとることができるので、二重結合の π 電子は O-C-O 全体にわたって存在することができます。このことを電子雲としてとらえると、電子雲が O-C-O 全体にわたって広がって分布し、密度も薄くなります。この電子雲は電子そのものであるので当然マイナスに荷電しています。したがって、その電子雲が密度が高くなるということは、その分内部に反発を抱えることになり、高エネルギーとなります。一方で、密度が低くなると内部に抱える反発も低減するため、低エネルギーとなります。このように共鳴構造をとらない場合には電子密度が高くなるため、高エネルギーとなる一方で、共鳴構造をとる場合には電子が存在できる場所が増えて、電子密度が低下するために低エネルギーとなります。
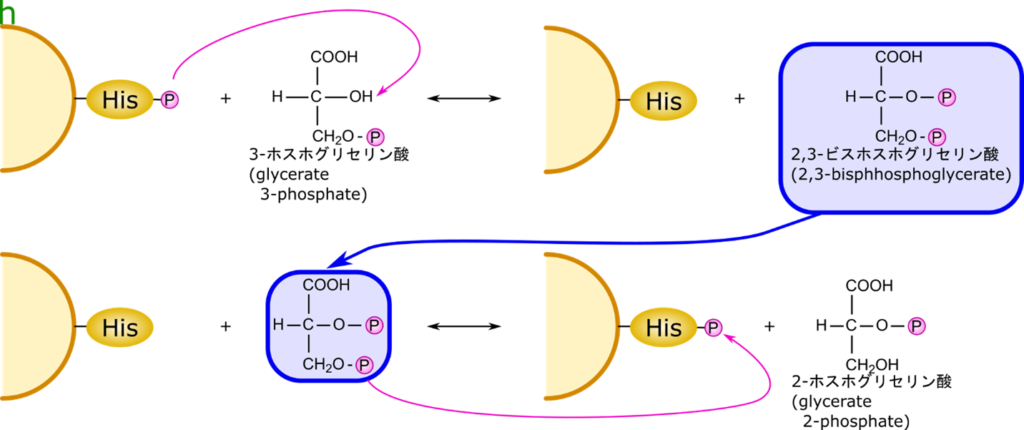
3-ホスホグリセリン酸から 2-ホスホグリセリン酸の反応は異性化ではなく転位反応
この反応は単純にリン酸基を移動させる反応ではなく、一度 2,3-ビスホスホグリセリン酸を経由して 2-ホスホグリセリン酸に変換される反応です。図8にこの反応の作用機序を記載しています。酵素上のヒスチジン残基にリン酸基が結合しています。このリン酸基が 3-ホスホグリセリン酸の 2 位の水酸基に転位させられます。その結果、2,3-ビスホスホグリセリン酸が生成されます。さらに 3 位のリン酸基がヒスチジンに引き渡されて、2-ホスホグリセリン酸が産生されます。ちなみに、酵素は初期状態に戻っていますね。
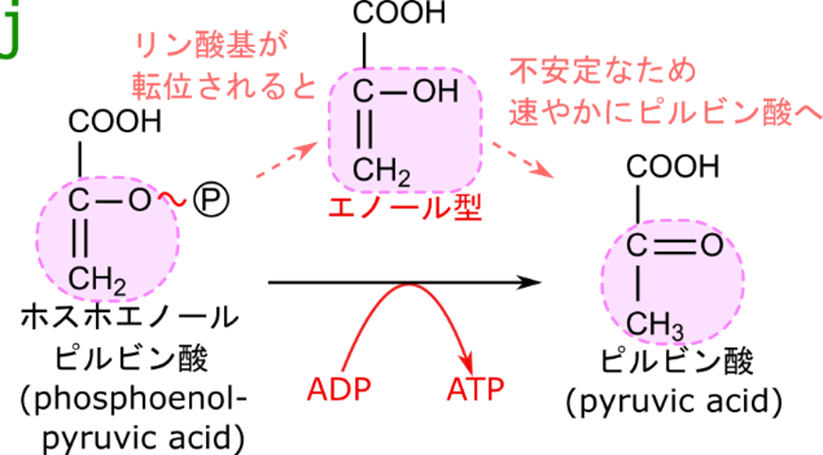
ホスホエノールピルビン酸からリン酸基とエネルギーを ADP へ引き渡す
最後にホスホエノールピルビン酸からピルビン酸への反応について解説します。この反応は脱リン酸することによりエネルギーを取り出す反応ですが、単純にリン酸基を ADP に渡している反応ではないことに気づくでしょうか?ホスホエノールピルビン酸からリン酸基をとってもピルビン酸にはなりません。ホスホエノールピルビン酸からリン酸基が除去されると、ピルビン酸のエノール型に変換されます。エノール型は極めて不安定であるため速やかにピルビン酸へ変換されます。したがって、この反応の反応産物はピルビン酸と ATP ということになります。この過程はケト-エノール互変異性の一例です。
先ほどエノール型は極めて不安定だということを言いました。一方でホスホエノールピルビン酸の構造に注目してください。この構造はエノール型を内包していますが、リン酸基が付いているためにピルビン酸への変換ができない状態になっています。リン酸基が ADP に渡されることにより水酸基がフリーになるのでピルビン酸への変換が速やかに行われ、安定化します。つまり、ホスホエノールピルビン酸はリン酸基のせいでケトンになって安定化することができないため(リン酸基をとることで安定化するため)、リン酸基の結合部分が高エネルギー結合になっています。したがって、脱リン酸化するとリン酸とエネルギーを ADP に渡して ATP を産生することができます。
練習問題
今回は解糖系について、特に反応メカニズムに焦点を当てて解説してきました。解糖系は進化的にきわめて古く、原核生物から真核生物に至るまで広く保存されている代謝経路です。そのため、解糖系は多くの代謝経路と密接に関連し、代謝全体を理解するうえでの基盤となる重要な経路と位置づけられます。
また解糖系の各反応には、ケト–エノール互変異性やアルドール反応など、糖化学や有機化学で学ぶ基本的な反応機構が数多く含まれています。解糖系を詳細に追うことは、生化学と有機化学を結び付けて理解するよい訓練にもなります。
本記事では、解糖系の化学的側面をできるだけ直感的に理解できるよう心がけて解説しました。ぜひ図や反応機構を見返しながら繰り返し読み、解糖系の反応メカニズムを確実に身につけてください。この理解は、今後学ぶTCA回路やエネルギー代謝の制御を理解するうえでも大きな助けとなるはずです。
参考文献
- 島原健三 (1991). 概説 生物化学. 三共出版. pp. 151-162
- Jeremy M. Berg, John L. Tymoczko, Gregory J. Gatto Jr., Lubert Stryer著、入村達郎、岡山博人、清水孝雄、中野徹訳 (2018). ストライヤー生化学 第8版. 東京化学同人. pp. 416-442
- John McMurry著、伊東椒、児玉三明訳 (2000). マクマリー有機化学 第4版. 東京化学同人. pp. 365-367, 546-551
- Paula Y. Bruice 著、大船泰史、香月勗、西郷和彦、富岡清訳 (2016). ブルース有機化学概説 第3版. 化学同人. pp. 678-729
関連記事
糖鎖の基本について
糖鎖の化学反応について
オリゴ糖・多糖について
解糖系について
- 解糖系(glycolysis)によって糖からエネルギーを取り出しATP を生成する
- 解糖系の各反応をエネルギー的観点から読み解く
- 解糖系で利用される糖はグルコースに限らない
- 解糖系の制御は三つの律速酵素に対する調節で行われる
TCA サイクルについて