ATPの合成は、ミトコンドリア内膜におけるプロトン勾配に依存しています。このプロセスは「化学浸透圧説(chemiosmotic hypothesis)」として知られており、ATP合成を説明する重要な仮説です。今回は、この化学浸透圧説の詳細、ATP合成の仕組み、そしてプロトン勾配がどのようにATP合成に寄与するかについて解説します。
Contents
化学浸透圧説(chemiosmotic hypothesis)とは?
プロトン勾配は、ミトコンドリア内膜での電子伝達によってプロトンが内膜外へ汲み出されることで形成されます(こちらの記事で詳細に説明しています)。この結果マトリックス側がマイナスに、細胞質側がプラスに帯電して電位差が生じます。この電位差は「膜電位」と呼ばれ、通常0.14V(ΔG゜’ = -5.2 kcal/mol)です。
プロトンは、膜電位によってマトリックス側に引き付けられ、その移動がATP合成酵素の駆動力となり、ATPが生成されます。この駆動力は「プロトン駆動力(proton-motive force)」と呼ばれ、pH勾配(プロトン濃度の違い)と電位勾配(電荷の差)によって成り立っています。
プロトン駆動力(Δp)
プロトン駆動力は、次の式で表されます:
Δp = ΔpH + Δψ
pH(プロトン濃度の対数)勾配は、プロトンの濃度差を解消する方向に動く力で、電位勾配はプロトンが電荷の差によって動く力です。
クリステによるプロトン勾配の効率化
ATP合成を行うミトコンドリア内膜は、ひだ状の構造(クリステ)を持っています。これにより、プロトン勾配を効率よく形成することができ、局所的にプロトン濃度を高めることが可能です。ATP合成酵素は二量体を形成し、それが集まってオリゴマーを形成することで、物理的に回転する ATP 合成酵素を安定化させると同時に、ひだ状のクリステを形成します。
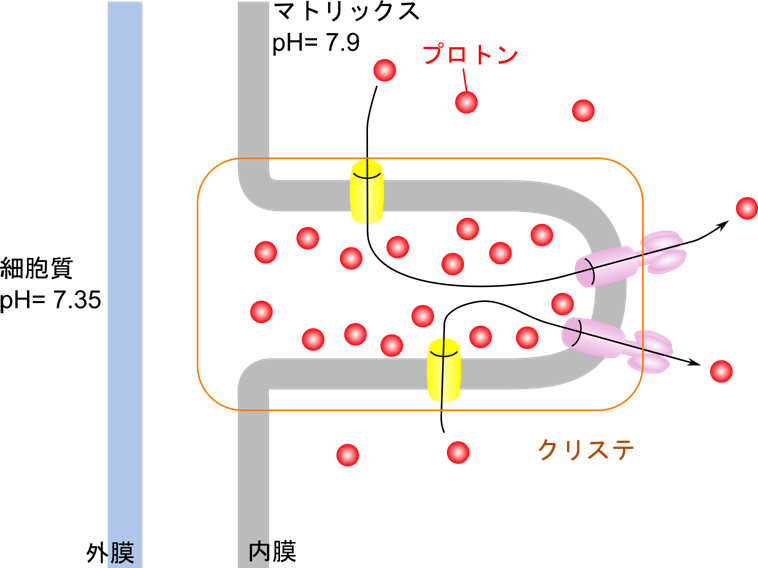
ATP 合成酵素は ATP 合成部分とプロトン移動によって回転する水車部分で成り立つ
ATP 合成酵素の構造
ATP合成酵素は、F0 部分とF1 部分に分かれています(図2)。F0 部分は、8~14個のcサブユニットが円形に並び、プロトンチャネル(プロトンの通り道)を形成しています。このプロトンチャネルは c リングと呼ばれています。F1 部分は、α サブユニットと β サブユニットが交互に並んだ六量体で、中心にγサブユニットがあります。プロトンがF0 部分を通過する際にF0 が回転し、これにより γ サブユニットも回転します。この回転がATP合成を駆動しますが、αおよびβサブユニットは回転せず、ATP生成の場となります。
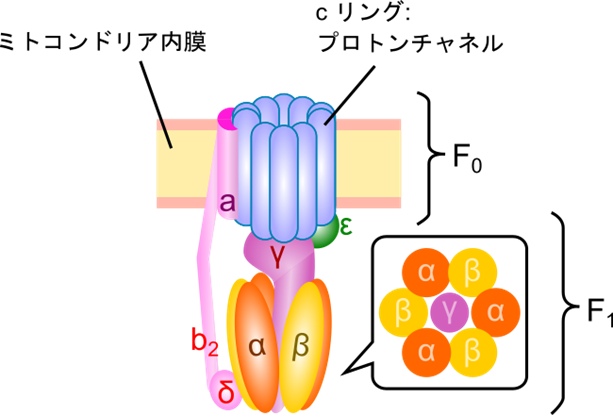
ATP 合成・放出の過程
ATP の産生は ADP に無機リン酸が脱水縮合することで行われます。この反応は無機リン酸のリンを ADP の酸素が攻撃することにより始まります。すると中間体を介して水が脱離して ATP となります。
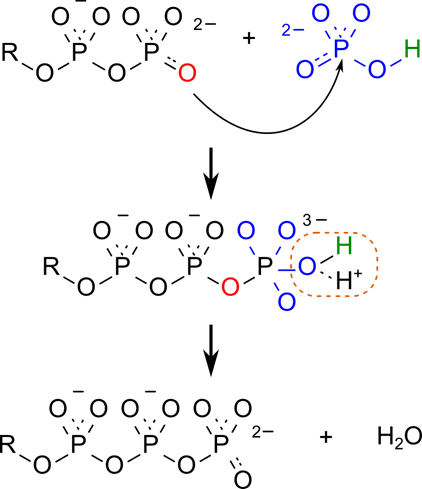
この反応を踏まえたうえで、ATP 合成の過程を見ていきましょう。ATP の合成は次の三つのステップで行われます。
- ADP と Pi(無機リン酸)が β サブユニットと結合します。
この結合は比較的緩いため、L(Loose)構造と呼ばれます。 - ATP が合成されます。
この結合は強いため、T(Tight)構造と呼ばれます。 - β サブユニットが開いた状態になり、ATP が放出されます。
この構造では開いた構造をとるため、O(Open)構造と呼ばれます
このステップの切り替わりは γ サブユニットの回転によりコントロールされています。図3で γ サブユニットの凸部分は β サブユニットを T 構造に、凹部分は β サブユニットを O 構造に、残りの部分は β サブユニットを L 構造にします。L 構造にされた β サブユニットではステップ1が実行されます。次に γ サブユニットが(反時計回りに)回転するとこの個所は T 構造になり、ステップ2が実行されます。さらに γ サブユニットが(反時計回りに)回転するとこの個所は O 構造になり、ステップ3が実行されます。このように γ サブユニットが回転することにより ATP 合成が進行するメカニズムは結合回転説(binding-change mechanism)として知られています。
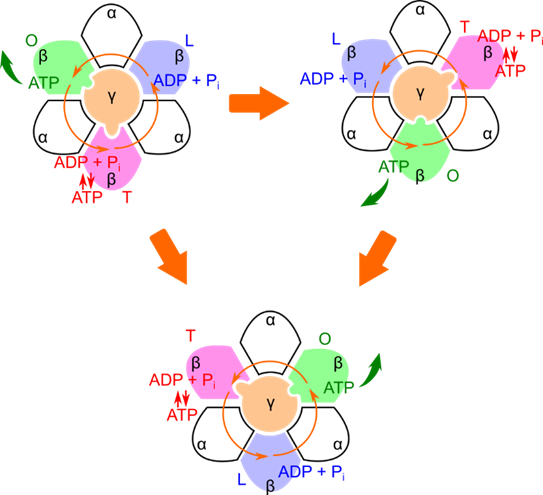
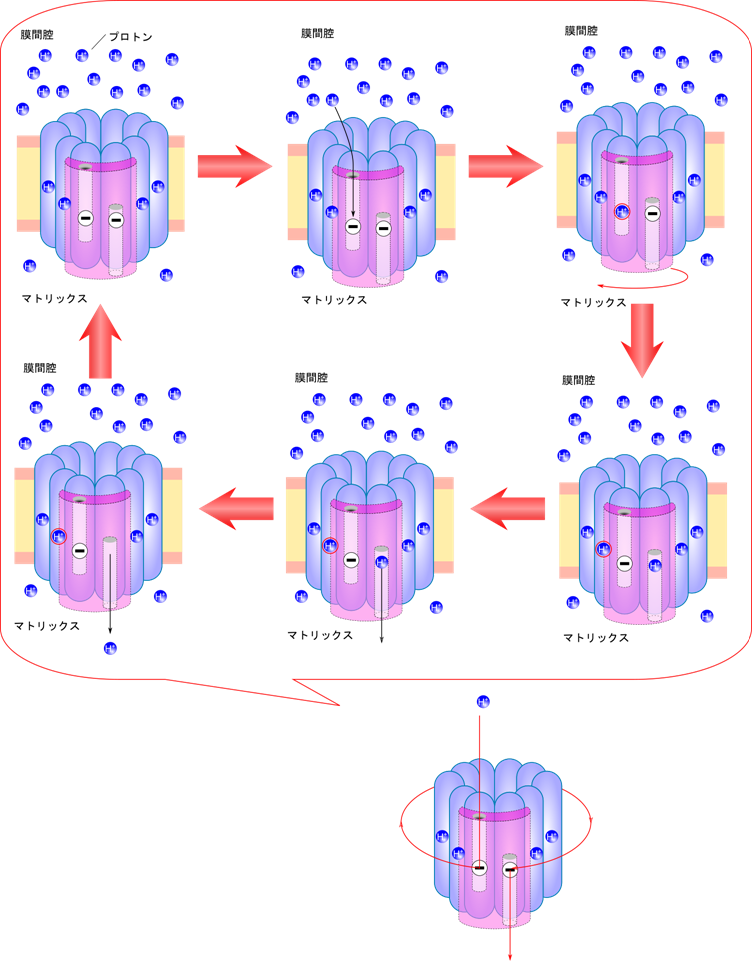
プロトンの流れによるATP合成酵素の回転
ATP合成酵素が回転する原動力は、c リングの回転です。では c リングはどのようにして回転するのでしょうか?a サブユニットには二つの半チャネルが存在しています。半チャネルは a サブユニットを完全に貫通していません。一つの半チャネルは細胞質側にもう一方はマトリックス側に開口部を向けています(図4)。c サブユニットの半チャネルの内部に当たる位置にグルタミン酸あるいはアスパラギン酸残基があり、プロトンを補足できるようになっています。細胞質側はプロトン濃度が高いので、プロトンは c サブユニットに捕捉され、c リングが回転します。この過程が繰り返されると c サブユニットは一周して a サブユニットのもう一方の半チャネルまで到達して、プロトン濃度が低いマトリックス側にプロトンを放出します。この過程は水力発電に似ています。水流のようにプロトンの流れが、水車のような ATP 合成酵素を回してエネルギー(ATP)を合成する仕組みとなっています。
酸化的リン酸化の制御機構
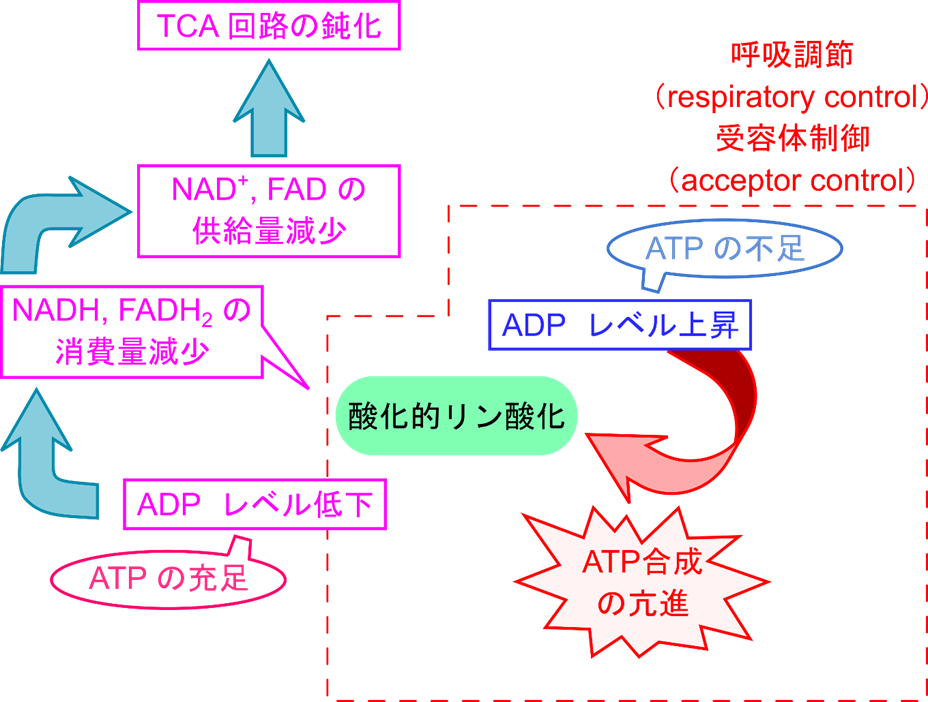
酸化的リン酸化は ADP レベルによってコントロールされている
ATP合成は、ADPの濃度に応じて制御されています。筋肉での運動時にはADP濃度が上昇し、それに伴い酸化的リン酸化速度は上昇します。これを「呼吸調整(respiratory control)」または「受容体制御(acceptor control)」と呼びます。一方、ADP濃度が低下すると、酸化的リン酸化によるNADHやFADH2の消費が減少することで、 NAD+ や FAD の産生量が低下し、TCA回路の機能も低下します。
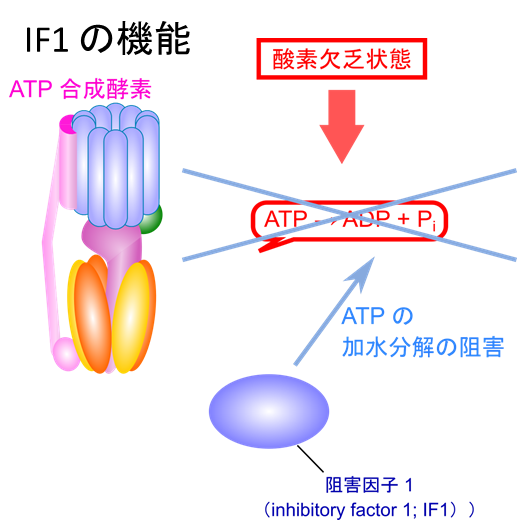
ATP合成酵素の逆回転の防止
酸素欠乏状態では、ATP合成酵素が逆回転し、ATPを無駄に消費してしまう可能性があります。この逆回転を防ぐため、阻害因子1(IF1)がATPの加水分解を阻止します。これにより、酸素欠乏時のATP消費を抑えるています。
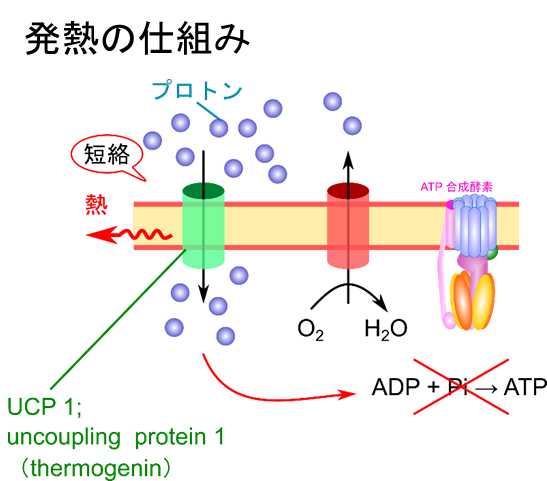
プロトン蓄電池をショートさせることで熱を発生させることができる
冬眠中の動物や新生児などでは、体温の維持のために脱共役タンパク質(uncoupling protein; UCP1、あるいはサーモゲニン; thermogenin)がプロトン勾配をショートさせることで、ATP合成を脱共役し、熱を発生させます。この反応は体温が低下した際に誘導され、アドレナリン作動性ホルモンによって脂肪酸が遊離し、UCP1を活性化します。
UCP1 は ATP-ADP トランスロカーゼに似たタンパク質で、UCP1 を通じてプロトンが流れた場合、本来 ATP 合成に利用されるはずのエネルギーは熱として放出されます。この反応は体内深部の体温が低下した場合に、アドレナリン作動性ホルモンが細胞質中の脂質粒子から脂肪酸を遊離させます。遊離した長鎖脂肪酸は UCP1 の細胞質表面に結合して UCP1 のプロトンの透過を誘導します。
今回は、ミトコンドリア内膜でのATP合成について解説しました。このプロセスは、自然界における水力発電と似ており、生物のエネルギー生成がいかに精巧にデザインされているかを示しています。
参考文献
- 島原健三 (1991). 概説 生物化学. 三共出版. pp. 139-150
- Jeremy M. Berg, John L. Tymoczko, Gregory J. Gatto Jr., Lubert Stryer著、入村達郎、岡山博人、清水孝雄、中野徹訳 (2018). ストライヤー生化学 第8版. 東京化学同人. pp. 485-519
関連記事
- プロトン勾配と酸化還元反応をエネルギー的な観点から解説:生体内のエネルギー流れ
- 細胞のエネルギー工場:ミトコンドリアとATP生成メカニズム
- 電子伝達系を徹底解説:プロトン汲み出しのメカニズムとは?
- 化学浸透圧説を徹底解説:プロトン勾配とATP合成の重要なつながり(←イマココ)
- NADHの電子はどう運ばれる?ミトコンドリア膜を越えた物質輸送を解説