グルコースレベルを一定に保つことはとても重要です。例えば脳の主要なエネルギー源はグルコースですし、赤血球に至ってはグルコースしか利用できません。特に飢餓状態の時はグリコーゲンなどの貯蔵分子も枯渇しているため、グルコースを生合成することに大きな意味が出てきます。今回はこのようなグルコースの生合成について解説していきます。
Contents
グルコースレベルを維持することの意義とは?グルコースのエネルギー源としての重要性
上述したように一部の組織ではグルコースが主要なエネルギーとして使用されています。ヒトの成人の脳が利用するグルコースは 120 g/day で、体全体が必要とするグルコース量は 160 g/day です。こうしてみてみると脳が利用するグルコースはヒトが利用する糖のほとんどを占めています。体液中のグルコース量は 20 g 程度であり、グリコーゲンから入手可能な量は 190 g にもなります。これは 1 日に必要なグルコース量 160 g よりも多いですね。ですので、グリコーゲンで貯蔵されているグルコース量は 1 日に必要なグルコース量に十分に見合う量となります。ところが飢餓状態に至ると、グリコーゲンによる貯蔵も底をつきますので、蓄えだけでは必要となるグルコース量を賄えなくなってきます。このような場合には糖新生がとても重要になってきます。
飢餓状態になると、足りないグルコースを補うため、乳酸、アミノ酸、グリセロールなどが適切に変換されて糖新生経路に流入します。乳酸やアミノ酸やピルビン酸又はオキサロ酢酸に変換されて、グリセロールはジヒドロキシアセトンリン酸に変換されてから糖新生に流入します。このようにしてグルコースレベルは常に維持されるわけですね。
単純な解糖系の逆経路のようで単純な逆経路ではない、糖新生の反応経路とは
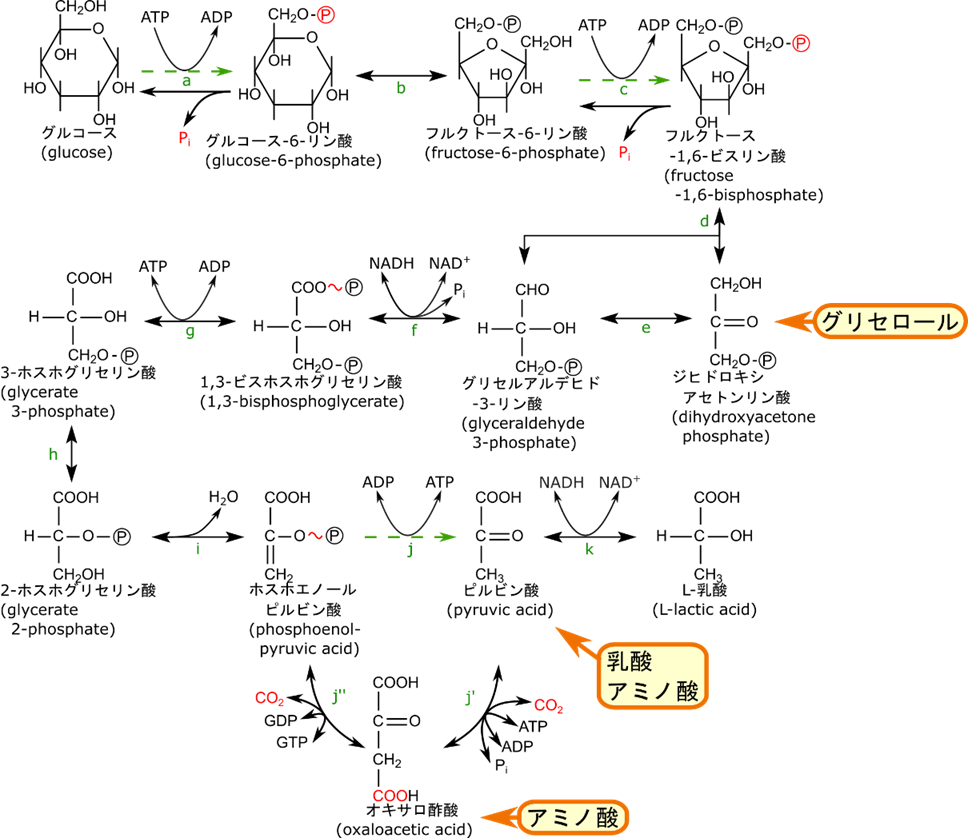
図1に糖新生経路を示しています。アミノ酸はオキサロ酢酸やピルビン酸に変換されてから流入します。一方で乳酸は無酸素条件下で解糖系が動くと産生されますが、乳酸デヒドロゲナーゼの作用でピルビン酸に変換されます。こうして産生されたピルビン酸が糖新生に流入ます。グリセロールについては以下の反応を介してジヒドロキシアセトンリン酸として糖新生に流入します。この反応はリン酸化とアルコールの酸化ですね。
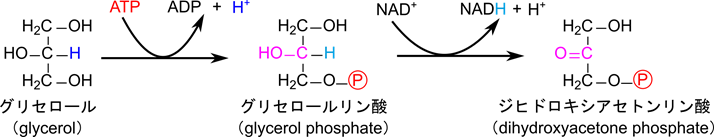
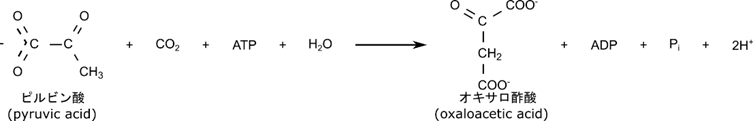
糖新生は基本的に解糖系の逆経路をたどります。ただし解糖系の反応 a, c, j は不可逆反応でした。そのため、この反応は逆方向にたどることができません。ですのでこれらの反応は異なる反応が起こります。反応 j ではまずピルビン酸がオキサロ酢酸に変換されます。この反応はミトコンドリアで行われます。
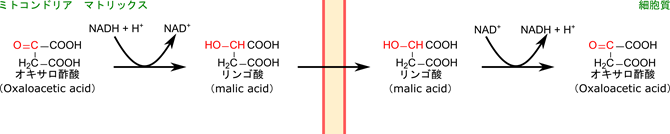
その後、産生されたオキサロ酢酸は細胞質に輸送されます。オキサロ酢酸はミトコンドリア膜を通過できないため、一度還元されて、リンゴ酸に変換されます。細胞質に輸送されると酸化されてオキサロ酢酸に変換されてのちの反応に利用されます。
最後にオキサロ酢酸がリン酸化されるとともに炭酸が脱離して、ホスホエノールピルビン酸に変換されます。
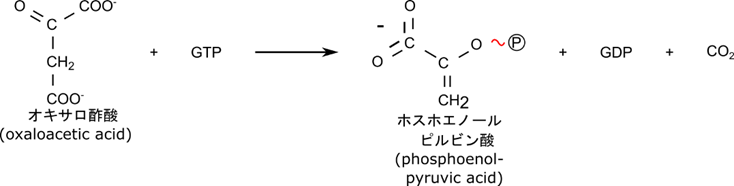
このようにして反応 j は迂回経路をたどることで不可逆反応をさかのぼることが可能になります。
さらに反応 c ではリン酸基を無機リン酸として除去します。この反応は解糖系の反応とは異なる反応で(逆反応なら ATP が産生されるはずですよね)、触媒する酵素はフルクトース-1,6-ビスリン酸ホスファターゼです。
反応 a はほとんどの臓器では起こらない反応になります。というのも、グルコース-6-リン酸は細胞外に拡散しないという利点があるため多くの臓器ではこちらの方が有利なためです。ただし、肝臓や(活性は弱いですが)腎臓などのように血中グルコース濃度を調節する機能を持つ組織ではグルコースまで変換されます。この場合はグルコース-6-リン酸のリン酸基が無機リン酸として除去されてグルコースに変換されます。
植物・細菌はグリオキシル酸回路によって脂質をグルコースへ変換できる
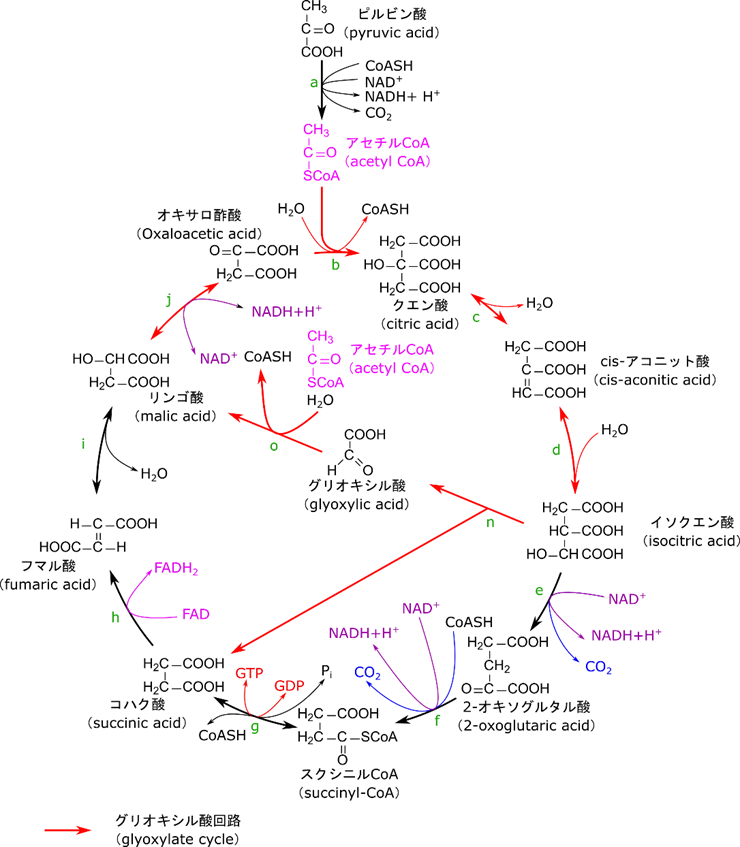
植物や一部の細菌はアセチル CoA からグルコースを合成する経路をもっています。前述の糖新生は各種生体分子からグルコースを生成する反応経路ですが、基本的に動物ではアセチル CoA からグルコースを生成することはできません。一方で(植物などが)グリオキシル酸回路(glyoxylate cycle)を利用すると、アセチル CoA からコハク酸を生成することができます。生成されたコハク酸はさらに糖新生とTCA 回路の組み合わせによってグルコースを合成することができます。
グリオキシル酸回路ではイソクエン酸までは TCA 回路と同様に反応が進行します。つまり、アセチル CoA をオキサロ酢酸が受け取ってクエン酸が生成され、クエン酸はアコニット酸を介してイソクエン酸に異性化します。ここでイソクエン酸(炭素数6)はグリオキシル酸(炭素数2)とコハク酸(炭素数4)に分解され、コハク酸が遊離します。グリオキシル酸はアセチル CoA からアセチル基を受け取ってリンゴ酸が生成されます。リンゴ酸は酸化されることでオキサロ酢酸になります。ここまでの反応ではアセチル CoA を二分子受け取って、コハク酸が生成されています。反応をまとめると
2アセチル CoA + NAD+ + 2H2O → コハク酸 + 2CoA + NADH + 2H+
ということになります。つまり、グリオキシル酸回路はアセチル CoA 2 分子からコハク酸を生成する反応経路と見ることができます。遊離したコハク酸は TCA 回路によってオキサロ酢酸に変換されますね。その後糖新生に入っていくことでグルコースの合成に利用されます。念のため確認しますが、この過程ではグリオキシル酸から生成されるオキサロ酢酸と、コハク酸から生成されるオキサロ酢酸がありますので、オキサロ酢酸が糖新生に供給されたとしても回路は回ります。さて、この反応はグリオキシソーム(glyoxysome)と呼ばれる細胞小器官で起こります。この回路はヒマワリやキュウリ、トウゴマなど油を多く含む趣旨で特に活発に行われます。この回路によって得られたグルコースなどの糖質が光合成ができるようになるまで植物に成長する力を与えます。
参考文献
- 島原健三 (1991). 概説 生物化学. 三共出版. pp. 151-162
- Jeremy M. Berg, John L. Tymoczko, Gregory J. Gatto Jr., Lubert Stryer著、入村達郎、岡山博人、清水孝雄、中野徹訳 (2018). ストライヤー生化学 第8版. 東京化学同人. pp. 416-442
関連記事
糖鎖の基本について
糖鎖の化学反応について
糖鎖の表記方法・三文字表記について
オリゴ糖・多糖について
解糖系について
- 解糖系(glycolysis)によって糖からエネルギーを取り出しATP を生成する
- 解糖系の各反応をエネルギー的観点から読み解く
- 解糖系で利用される糖はグルコースに限らない
- 解糖系の制御は三つの律速酵素に対する調節で行われる
TCA サイクルについて
糖質合成について