解糖系は三つの不可逆反応で調節を受けています。この反応を触媒する酵素は解糖系の速度を決定するので “律速酵素” と呼ばれます。ところで解糖系は単なるエネルギーの製造装置というわけではありません。様々な代謝経路に前駆体を供給する役割も担っています。解糖系の制御は他の代謝経路への枝分かれとも密接にかかわっています。というわけで今回は解糖系の制御と他の代謝経路との関連を解説していこうと思います。
Contents
解糖系は極めてち密に制御を受けている
解糖系の制御を考える上で重要なことは、対象としている解糖系は筋肉(末梢)の解糖系か肝臓の解糖系かを分けて考えることです。これは解糖系のもつ意義が筋肉では主にエネルギー生産なのに対して、肝臓では種々の代謝の材料を提供する役割や血糖(血中糖濃度)の調整にあるという違いによります。したがってどういうときに系を止めるべきかも筋肉(末梢)と肝臓で異なります。これに伴って制御系の働くメカニズムが異なってきます。
制御は不可逆反応のステップで行われる
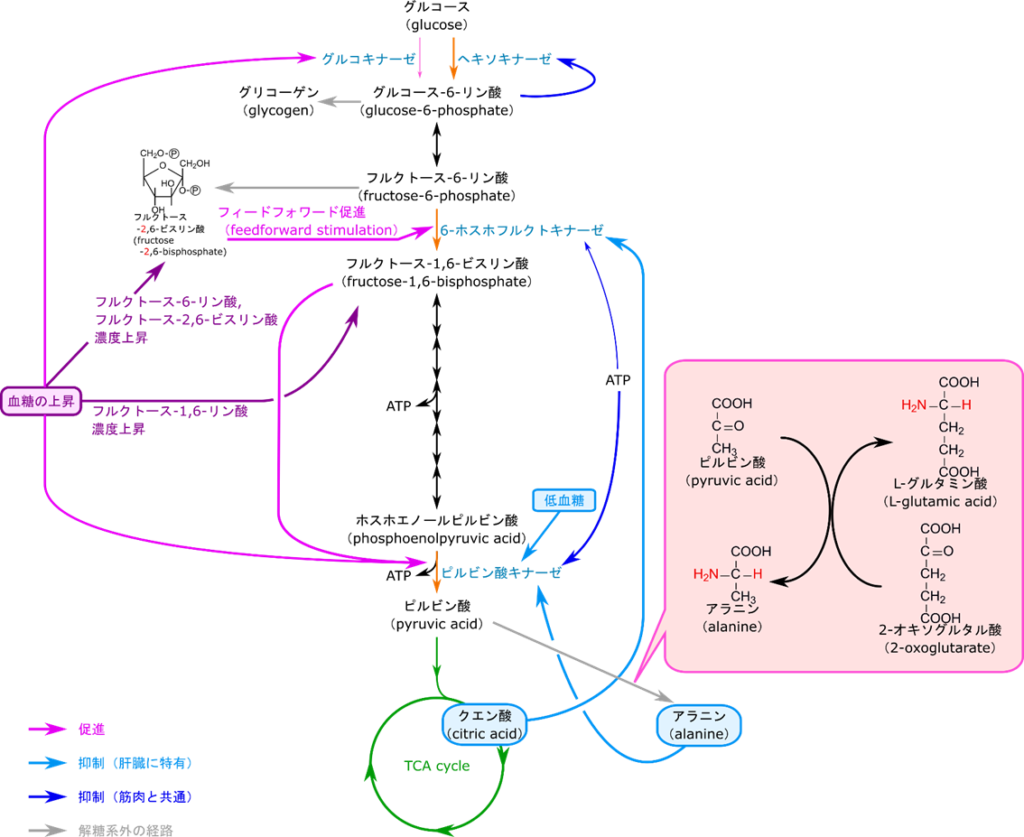
解糖系の制御は不可逆反応の段階に対して行われます。解糖系の反応の中で不可逆反応は図6で反応 a, c, j の三段階です。これらの反応はストップすると完全に反応が停止してくれるため、効率よく制御ができるステップです。さて、反応 a はヘキソキナーゼが触媒しており、反応 c は 6-フルクトキナーゼが、反応 j はピルビン酸キナーゼが触媒しています。したがって、解糖系の制御を考える場合にはヘキソース、6-フルクトキナーゼ、ピルビン酸キナーゼの三つの酵素の制御を考えることになります。それでは以降、制御の詳しい内容を見ていきましょう。
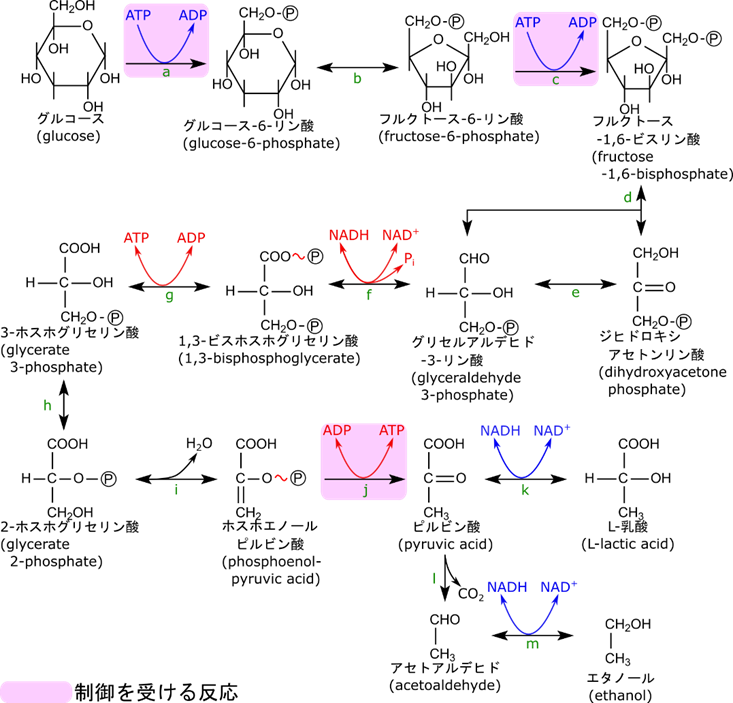
筋肉(末梢)における解糖系の制御。エネルギー生産の調節
ヘキソキナーゼの制御はフィードバック阻害によって行われる
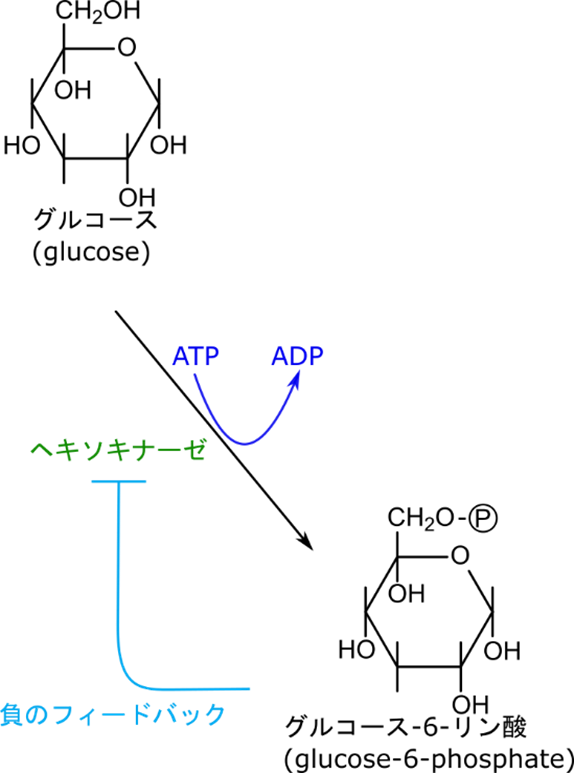
ヘキソキナーゼはグルコースをリン酸化してグルコース-6-リン酸にする反応を触媒します。この際 ATP を消費します。この反応の制御は単純なフィードバック阻害です。フィードバック阻害とは生成物が酵素の機能を阻害することを言います。戻るようにして阻害をするためフィードバック阻害と呼ばれます。今回の場合はグルコース-6-リン酸がヘキソキナーゼの機能を阻害します。
それでは経路の中での立ち位置から考察してみましょう。この反応の生成物であるグルコース-6-リン酸は次の反応でフルクトース-6-リン酸に変換されます。実はフルクトース-6-リン酸を作る反応は ATP レベルによって阻害がかかる反応です。ですのでエネルギーが過多になった場合反応がストップしてしましいます。エネルギーが充足している状態ではフルクトース-6-リン酸濃度が上昇します。この反応のグルコース-6-リン酸とフルクトース-6-リン酸は平衡を作っているのでフルクトース-6-リン酸が過剰な場合にはその一部がグルコース-6-リン酸に変換されます。その結果グルコース-6-リン酸レベルも上昇して、ヘキソキナーゼが阻害を受けるようになります。
6-フルクトキナーゼの制御は ATP と AMP によって調節される
二つ目の制御ポイントは 6-フルクトキナーゼに対する制御です。この制御は説明の順番上二番目に解説していますが、実はこの制御が解糖系全体の制御の起点になるかなり大事な制御です。では見ていきましょう。
6-フルクトキナーゼの制御には ATP と AMP が大きく関わっています。このことを端的に示す数値として ATP/AMP 比を利用します。この数値は ATP が大きくなると大きくなり、AMP が多くなると小さくなります。
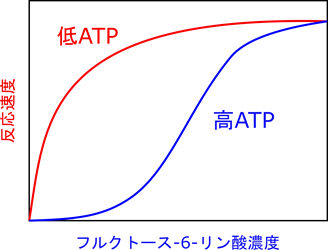
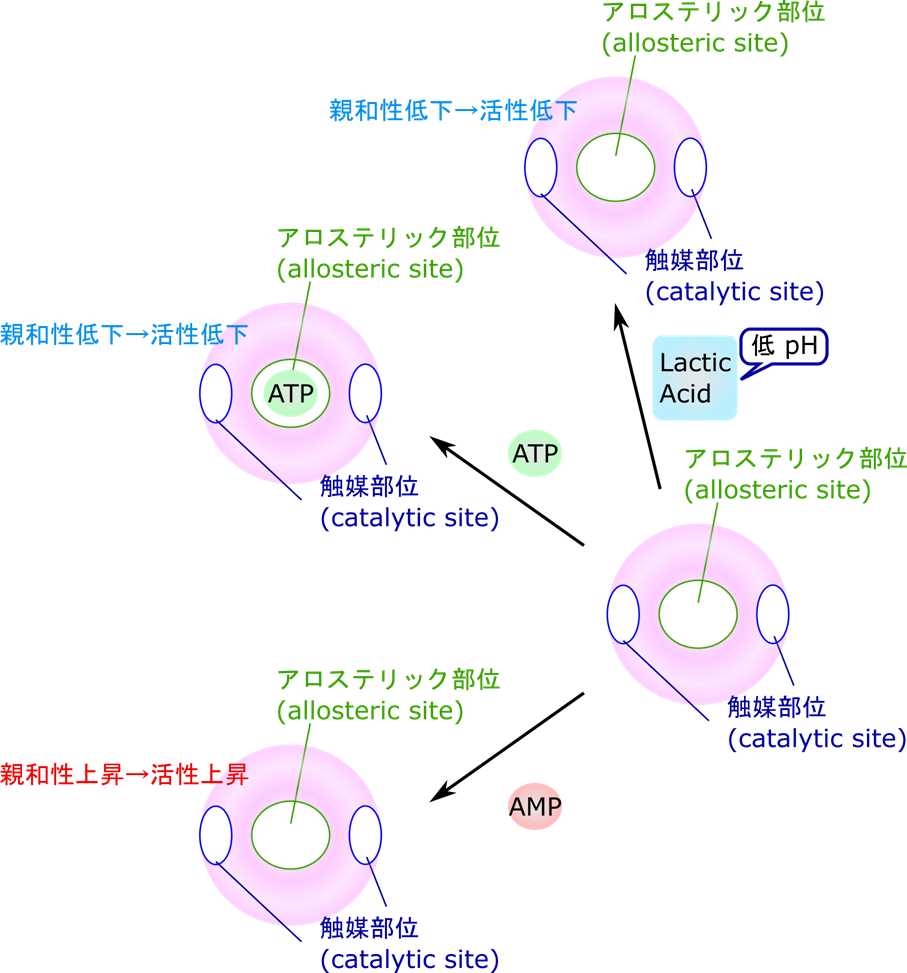
6-フルクトキナーゼには活性を発揮する触媒部位とは別に ATP が結合するアロステリック部位(allosteric site)が存在します。この部位が制御のポイントになります。もし、エネルギーが充足すると ATP がたくさん存在します(高ATP/AMP 状態)。すると ATP がアロステリック部位に結合します。その結果、触媒部位の基質との親和性が低下します。活性が十分に発揮される条件下ではフルクトース-6-リン酸濃度(基質濃度)と反応速度の関係を調べると右上図の赤線のようになります。一方で基質との親和性が低下すると基質濃度と反応速度の関係は青線のようなシグモイド関数(S字状の関数)になります。赤線の場合は(特に基質の低濃度域で)少しの濃度上昇だけで鋭敏に反応して反応速度が大きく上昇します。ところが青線の場合には、なかなか速度が上昇せず、基質濃度をだいぶ上昇させないと反応速度が上昇しません。このように、赤線から青線のような反応速度のシフトが起こると反応速度は大幅に減少してしまいます。さらに、解糖系が進むと乳酸がたくさん産生されていきます。すると pH が低下してしまいます。この低 pH は ATP の作用を増大させてさらに反応速度を低下させます。一方で、ATP/AMP 比が低下する(AMP 量が上昇する)と、AMP が ATP の作用を阻害して酵素活性は上昇します。このような制御の方式をアロステリック制御(allosteric regulation)といいます。
ところで、正の制御を司るのは AMP でしたね。なぜ ADP ではなく AMP なのでしょうか?ATP が急激に消費されると以下の反応を介して AMP が産生されます。
ADP + ADP ⇆ ATP + AMP
このようにして ADP が再利用されて ATP が供給されます。この時 AMP が産生されますが、この AMP は低エネルギーであることの合図になります。ところで、ATP と ADP、AMP の総量は短い時間で考えた場合一定になります。さらに ATP、ADP、AMP の濃度は [ATP]>[ADP]>[AMP] という関係にあります。この二つの事実から ATP 濃度が少し変化するだけで AMP 変化は大きく変動することになります。そのため、AMP に制御を頼ることで感度が高くなり、より厳密な制御が可能になります。
ピルビン酸キナーゼは ATP とフルクトース-1,6-ビスリン酸によって調節される
ピルビン酸キナーゼは ATP によるフィードバック阻害を受けます。エネルギーが充足した場合、ATP が豊富に存在することになります。こうして産生された ATP がアロステリックに阻害してピルビン酸キナーゼの活性が低下します。一方で、フルクトース-1,6-ビスリン酸はピルビン酸キナーゼが触媒する反応のずっと上流にある物質です。この物質がたくさん存在するということは系を促進しないといけない状況となりますね。なのでフルクトース-1,6-ビスリン酸はピルビン酸キナーゼの活性を増大させます。
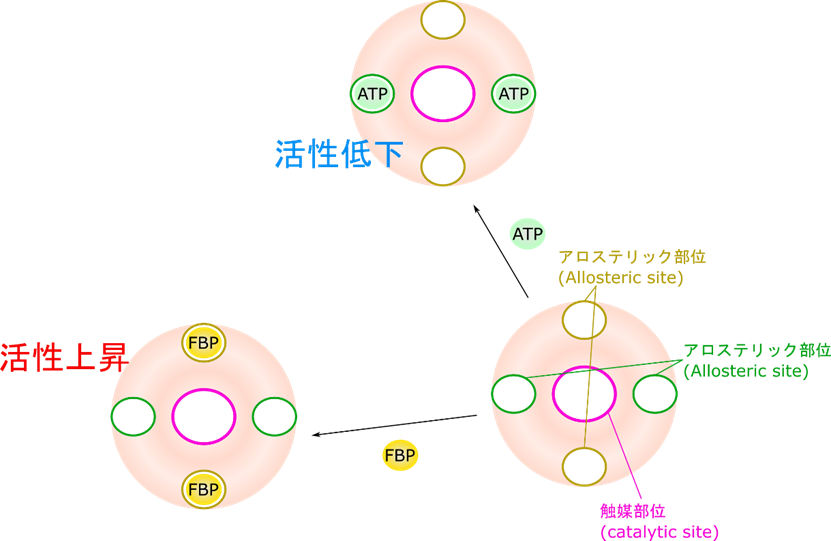
FBP: フルクトース-1,6-ビスリン酸
筋肉における解糖系の抑制は ATP による制御に始まり、促進は AMP による促進に始まる
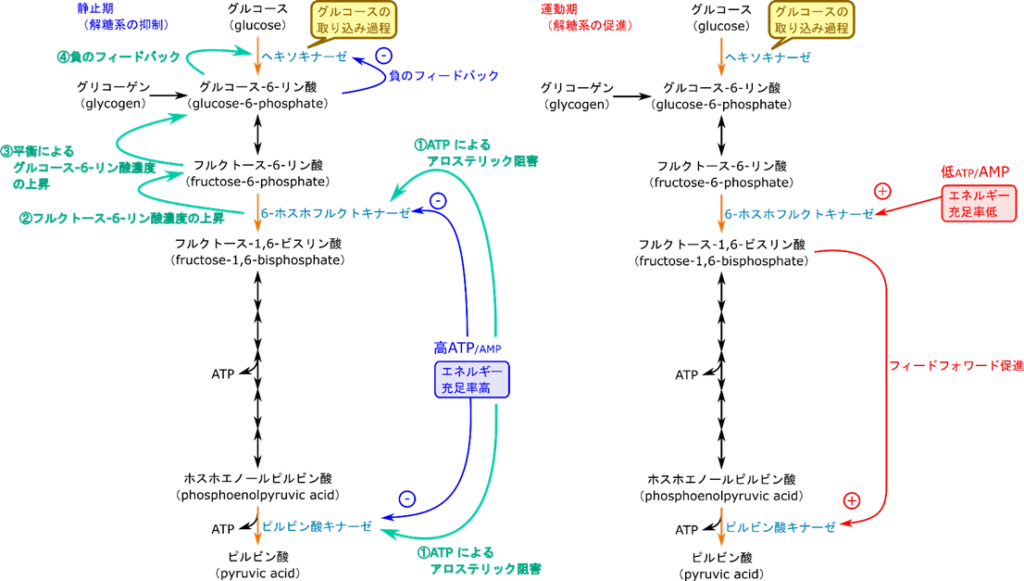
これまで、制御の各論を見てきました。これらをまとめて議論しようと思います。まず、エネルギーが充足した静止期には解糖系の抑制が行われます。この制御ではまず高 ATP/AMP 比が 6-ホスホフルクトキナーゼを抑制するとともに、ピルビン酸キナーゼを抑制します。6-ホスホフルクトキナーゼの活性が低下するとフルクトース-6-リン酸を消費する反応の速度が低下するためフルクトース-6-リン酸の濃度が上昇します。グルコース-6-リン酸とフルクトース-6-リン酸の異性化は平衡反応なので、フルクトース-6-リン酸の濃度が上昇すると一部が戻ってグルコース-6-リン酸に変換されます。するとグルコース-6-リン酸の濃度が上昇するのでヘキソキナーゼのフィードバック阻害が強まります。これらの制御が強く働くため解糖系は全体として抑制されます(図10左)。
一方で運動期にはエネルギーが要求されますので解糖系が促進されます。そうするとエネルギーが不足しますので ATP/AMP 比が低下します。低 ATP/AMP が 6-ホスホフルクトキナーゼの活性を促進します。すると 6-ホスホフルクトキナーゼが触媒する反応の速度が増大するのでフルクトース-1,6-ビスリン酸濃度が上昇します。このフルクトース-1,6-ビスリン酸がピルビン酸キナーゼの活性を増大させます。こうして解糖系は全体として促進されます(図10右)。
このように解糖系の制御はそれぞれがバラバラに行われるのではなく、全体が連携して行われます。ところで静止期、運動期ともに 6-ホスホフルクトキナーゼが調節の起点になっていることに気づくでしょうか?この点で 6-ホスホフルクトキナーゼは解糖系のペースメーカー的な役割を担っています。とてもよくできた仕組みですね。
肝臓解糖系の制御。血糖レベルと前駆体供給の調整
肝臓におけるグルコース取り込みはヘキソキナーゼとグルコキナーゼによって触媒される
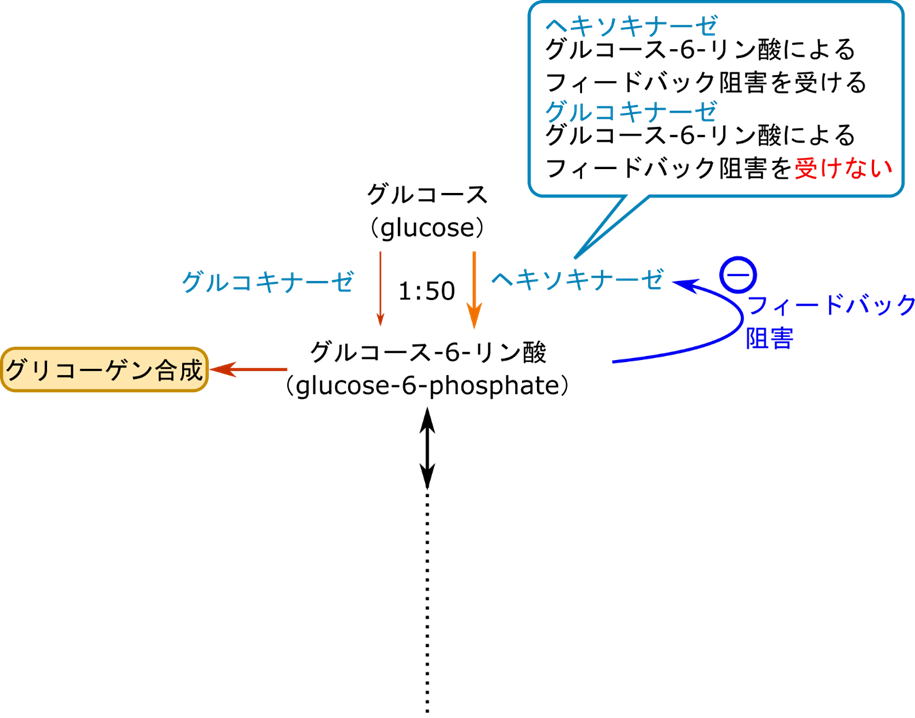
解糖系の最初の段階はヘキソキナーゼによるグルコースのリン酸化でしたね。この過程は血中から糖を取り込む過程でもあります。肝臓におけるこの過程の意味は解糖系の第一段階というだけでなく血糖値の調整やグリコーゲン合成のためのグルコース-6-リン酸の供給という意味があります。もし、高 ATP/AMP になった場合、ヘキソキナーゼはグルコース-6-リン酸のフィードバック阻害を受けてしまいます。そうするとグリコーゲンの合成のためのグルコース-6-リン酸供給まで停止してしまいます。ですのでヘキソキナーゼとは別にグルコース-6-リン酸の阻害を受けないグルコキナーゼによるグルコースのリン酸化も機能しています。二つも同じ機能を持つものが働いていると、供給が過剰になって制御がうまく機能しなさそうですよね。ですが、実際にはヘキソキナーゼとグルコキナーゼでは触媒する反応の反応速度が全く違います(ヘキソキナーゼ:グルコキナーゼ=50:1 程度)。その結果、ヘキソキナーゼが機能している間はグルコキナーゼ活性は影響しなくなります。一方でヘキソキナーゼの機能が低下してくるとグルコキナーゼの機能によりグルコース-6-リン酸の供給は維持されるという仕組みになっています。
肝臓の 6-ホスホフルクトキナーゼの調節は前駆体供給の調整と血糖値調整の意味がある
解糖系には TCA 回路に前駆体を供給する役割もあります。解糖系の最終産物であるピルビン酸はアセチルCoA としてTCA 回路に供給され、クエン酸の合成に使用されます。クエン酸がたくさん存在するようになると TCA 回路にもはやクエン酸の供給は必要なくなります。このようなシチュエーションではクエン酸は 6-ホスホフルクトキナーゼに対する ATP の作用を増大させてその機能を阻害し、ピルビン酸の供給を停止させます(図12の左側)。
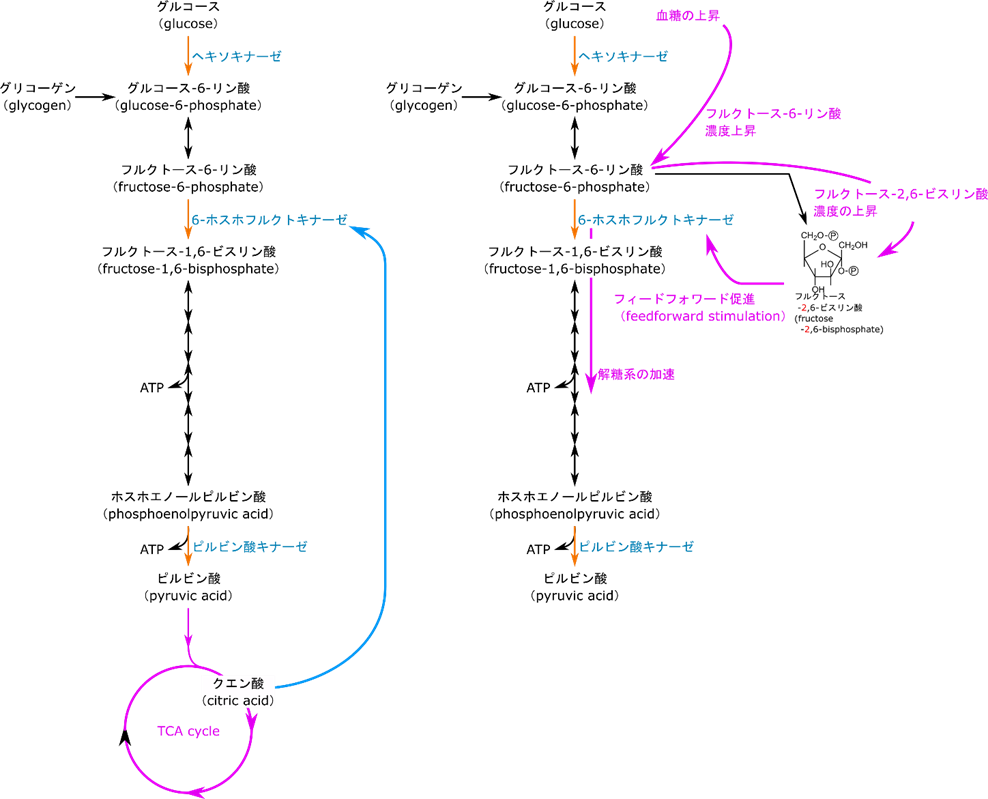
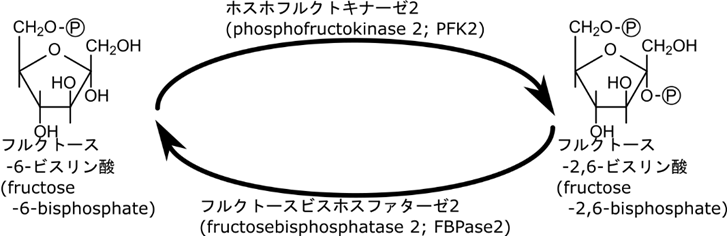
一方で血糖値が高いときは、グルコースがどんどん供給されるのでフルクトース-6-リン酸濃度が上昇します。このようなシチュエーションでは 6-ホスホフルクトキナーゼの機能を上昇させて処理しないといけません。フルクトース-6-リン酸がホスホフルクトキナーゼ2の作用によってフルクトース-2,6-ビスリン酸(フルクトース-1,6-ビスリン酸ではありません)が産生されます。一方で低血糖になるとフルクトース-2,6-ビスリン酸はフルクトースビスホスファターゼ2の作用によってフルクトース-6-リン酸に変換されます(図13)。このフルクトース-2,6-ビスリン酸は 6-ホスホフルクトキナーゼとフルクトース-6-リン酸の親和性を上昇させて6-ホスホフルクトキナーゼの活性を増大させます。このフィードフォワード促進のキー分子はフルクトース-2,6-ビスリン酸ですね。この分子の生合成はどのようにして制御されているのでしょうか?次はその制御についてみていきましょう。
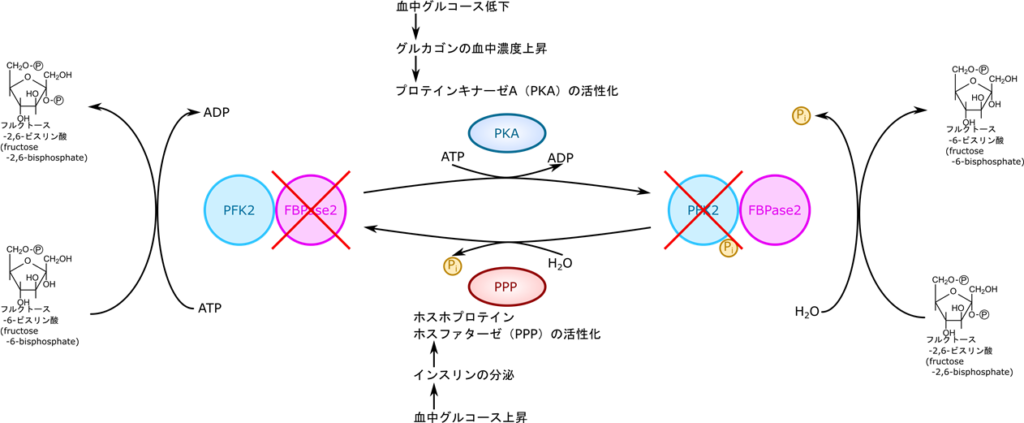
フルクトース-2,6-ビスリン酸の合成はホスホフルクトキナーゼ2とフルクトースビスホスファターゼ2の作用にって合成されたり、逆にフルクトース-6-リン酸に変換されたりします。このように、ホスホフルクトキナーゼ2とフルクトースビスホスファターゼ2は全く真逆の作用を及ぼしますが、実際には一つの分子この相反する作用を担っています。この分子にはキナーゼの活性を担うキナーゼ部位とフォスファターゼの活性を担うフォスファターゼ部位があります。このままではリン酸化するものと脱リン酸化が同時に機能してしまい、うまく制御ができなさそうですね。ですのでそれぞれの部位のスイッチを ON/OFF する機構が存在します。高血糖の場合は高フルクトース-6-リン酸状態になるわけですからフルクトース-2,6-ビスリン酸を産生して 6-ホスホフルクトキナーゼの作用を増大させなければならないですね。この時はインスリン(情報伝達物質であるホルモンの一種です)の作用でホスホプロテインホスファターゼが活性化されキナーゼ部分が脱リン酸化されてキナーゼ部分が “ON” になります。すると上記の通りフルクトース-2,6-ビスリン酸が合成されます。一方で低血糖の場合はグルカゴン(ホルモンの一種)の血中濃度が上昇します。その結果プロテインキナーゼ A が活性化されて、このプロテインキナーゼ A の作用によって、キナーゼ部分がリン酸化されて機能が “OFF” にされ、ホスファターゼ部分が “ON” になります。すると、フルクトース-2,6-ビスリン酸がフルクトース-6-リン酸に変換されます。このように、肝臓における 6-ホスホフルクトキナーゼの活性調整は血糖値の高低によって制御されています。
肝臓解糖系のピルビン酸キナーゼは血糖値の高低によって制御される
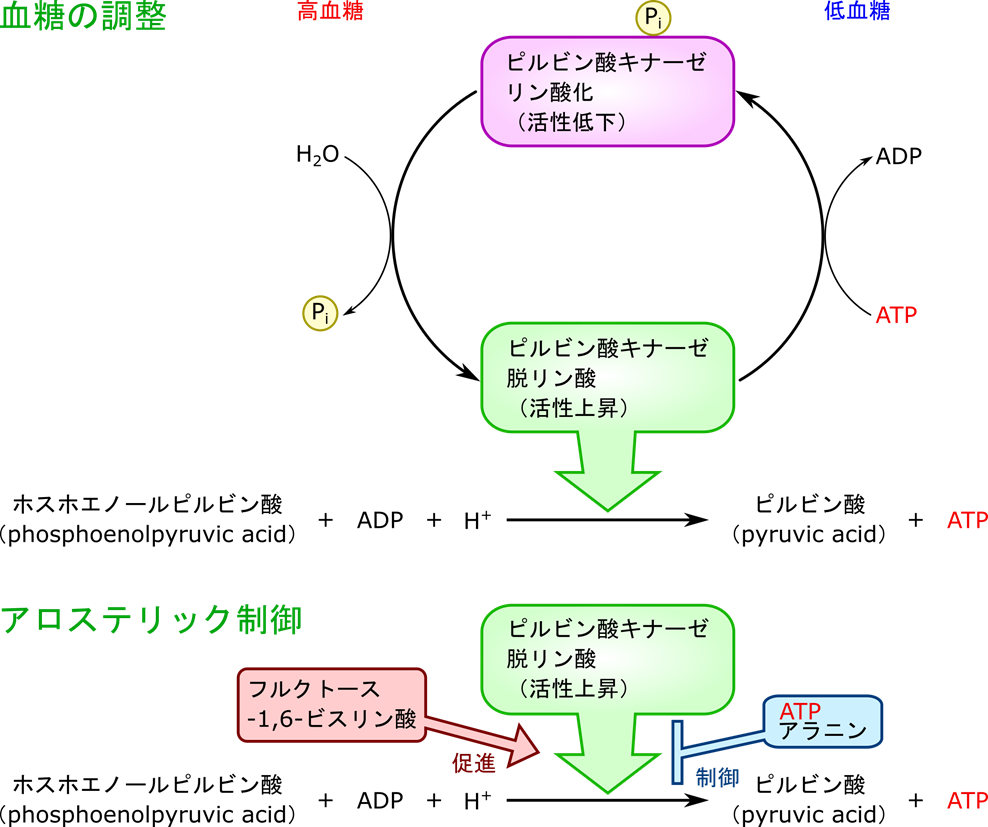
ピルビン酸キナーゼには筋肉で機能する M 型(Muscle)と肝臓で機能する L 型(Liver)が存在します。このため制御メカニズムもそれぞれで異なります。L 型は高血糖の場合、脱リン酸化を受けて、ピルビン酸キナーゼの活性は上昇します。一方で低血糖ではグルカゴンの作用によりリン酸化されてピルビン酸キナーゼの活性は低下します。
一方で M 型ピルビン酸キナーゼ(筋肉)と同様、フルクトース-1,6-ビスリン酸によるフィードフォワード促進も受けます。一方で、ピルビン酸はアラニンの前駆体でもあります。図16に示したようにピルビン酸は L-グルタミン酸と共役してアラニンに変換されます。ですので、L 型ピルビン酸キナーゼはアラニンによるフィードバック阻害を受けることによりアラニン合成の前駆体供給調整も行われます。
血糖値による調整(上)とアロステリックな制御(下)
肝臓における解糖系の制御は前駆体供給と血糖値を調整する
肝臓における解糖系の制御をまとめてみました(図16)。グルコース-6-リン酸の合成過程では筋肉と同様ヘキソキナーゼも機能していますが、グルコキナーゼも同時に機能しています。グルコキナーゼはヘキソキナーゼがフィードバック阻害によって停止した場合においても継続して機能し続けます。このことはグリコーゲンの合成のためにグルコース-6-リン酸を供給する意味があります。6-ホスホフルクトキナーゼの活性はクエン酸によって抑制されます。この制御は TCA 回路に対するアセチル CoA の供給調整の意味があります。さらにピルビン酸キナーゼはアラニンによる阻害を受けますが、このこともピルビン酸の供給調整の意味があります。以上のように肝臓の解糖系では様々な生体分子の生合成の前駆体を供給する役割を担っています。そのため、肝臓の解糖系の制御は他の代謝経路の前駆体供給調整というと側面があります。
一方で高血糖下ではフルクトース-6-リン酸がフルクトース-2,6-ビスリン酸に変換されて 6-ホスホフルクトキナーゼがフィードフォワード促進を受けます。一方で低血糖下ではフルクトース-2,6-ビスリン酸がフルクトース-6-リン酸に変換されてこのフィードフォワード促進はストップします。さらにピルビン酸キナーゼの制御は低血糖だと抑制され、高血糖だと促進されます。このように肝臓の解糖系は血糖値の調整の役割も担っています。
今回はグルコース以外の糖の利用と解糖系の制御についてまとめてみました。特に解糖系の制御には筋肉の解糖系と肝臓の解糖系の生理学的な意義の違いが大きく影響します。その観点から知識を整理していただけると理解しやすいのではないかと思います。最後まで読んでいただいてありがとうございます。
参考文献
- 島原健三 (1991). 概説 生物化学. 三共出版. pp. 151-162
- Jeremy M. Berg, John L. Tymoczko, Gregory J. Gatto Jr., Lubert Stryer著、入村達郎、岡山博人、清水孝雄、中野徹訳 (2018). ストライヤー生化学 第8版. 東京化学同人. pp. 416-442
- John McMurry著、伊東椒、児玉三明訳 (2000). マクマリー有機化学 第4版. 東京化学同人. pp. 365-367, 546-551 Paula Y. Bruice 著、大船泰史、香月勗、西郷和彦、富岡清訳 (2016). ブルース有機化学概説 第3版. 化学同人. pp. 678-729
関連記事
糖鎖の基本について
糖鎖の化学反応について
オリゴ糖・多糖について
解糖系について
- 解糖系(glycolysis)によって糖からエネルギーを取り出しATP を生成する
- 解糖系の各反応をエネルギー的観点から読み解く
- 解糖系で利用される糖はグルコースに限らない
- 解糖系の制御は三つの律速酵素に対する調節で行われる
TCA サイクルについて