ミトコンドリア内膜は基本的に物質の通過を許しません。ですが、外部で産生された NADH などミトコンドリア内で処理されなければならない物質はどのようにしてミトコンドリア内に運ばれるのでしょうか?今回はミトコンドリア膜をまたぐ物質移動のメカニズムについて解説します。
Contents
解糖系で産生された NADH は様々なシャトル分子と共役することで電子をミトコンドリア内に伝達する
解糖系のグリセルアルデヒド-3-リン酸の酸化により NADH が産生されます。この NADH が再度解糖系で使用されるためには NAD+ に再生される必要があります。ところが NADH はミトコンドリア内膜を透過できません。では、どのようにして NADH は NAD+ に再生され、電子伝達系に寄与するのでしょうか?
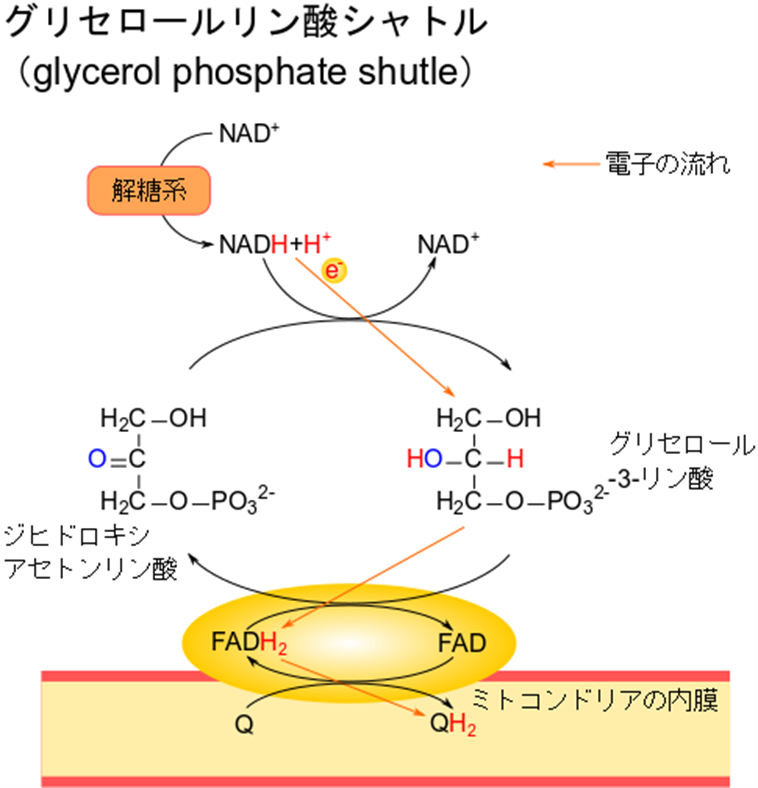
グリセロールリン酸シャトルによる電子の伝達
NADH はジヒドロキシアセトンリン酸に電子を渡すことでグリセロール-3-リン酸を産生します。その後、グリセロール-3-リン酸は FAD に電子を渡して酸化され、ジヒドロキシアセトンリン酸に戻ります。一方 FAD は FADH2 に変換されますが、FADH2 は酸化型ユビキノン(Q)に電子を渡して還元型ユビキノン(QH2)を産生します。この還元型ユビキノンは電子伝達系に供給されて、エネルギー生産に寄与します。解糖系の NADH はいきなり還元型ユビキノンから合流することになります。この経路では NADH 自体は輸送されませんが、電子が伝達されることで NADH が輸送されるのと同様の効果をもたらします。
この経路は FADH2 が電子伝達系で処理される経路と同様です。ですので、NADH であっても産生される ATP の分子数は FADH2 の時と同様で 2 分子となりますので、注意が必要です(NADH と FADH2 が何分子の ATP を生成するかはこちら)。この経路は筋肉で特に顕著で、グリセロール-3-リン酸とジヒドロキシアセトンリン酸を行ったり来たりするので、グリセロールリン酸シャトル(glycerol phosphate shutle)と呼ばれています。
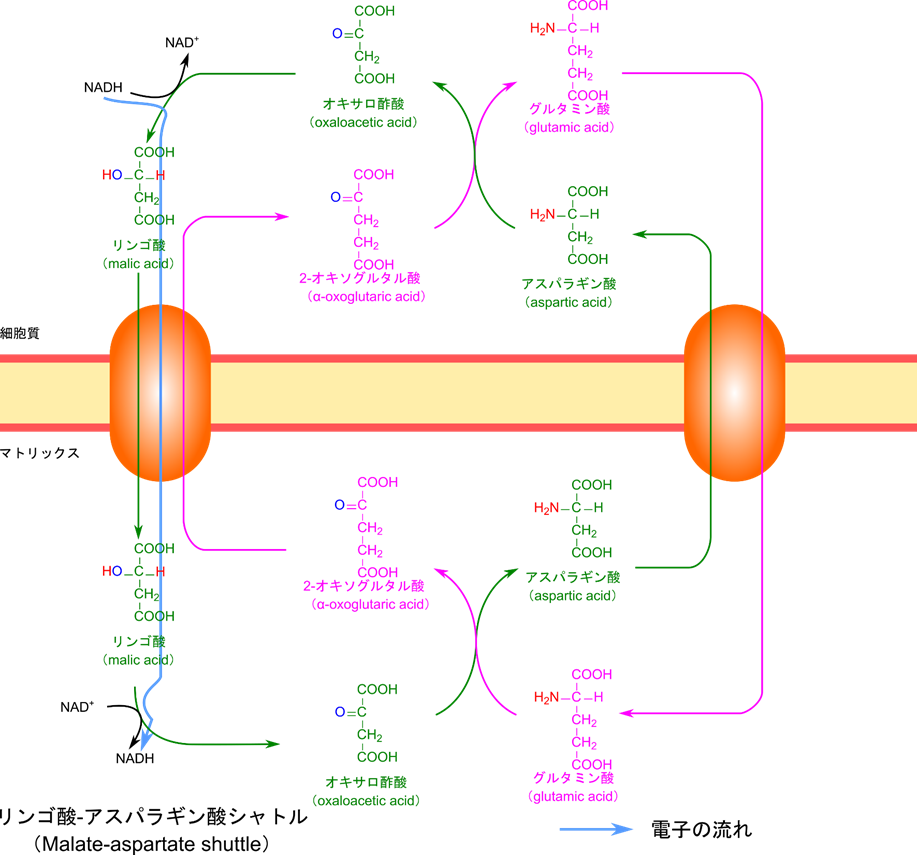
リンゴ酸-アスパラギン酸シャトルを介して NADH の電子がミトコンドリア内の NAD+ に伝達される
心臓や肝臓では、NADH は異なるメカニズムで処理されます。このメカニズムを図2に示しています。NADH はオキサロ酢酸に電子を渡して還元し、リンゴ酸を産生します。リンゴ酸は 2-オキソグルタル酸と入れ替わりにミトコンドリア内膜を通過し、NAD+ に電子を渡すことで酸化されオキサロ酢酸に変換されます。この際に NADH が産生されて、電子伝達系に供給されます。産生されたオキサロ酢酸は、グルタミン酸からアミノ基をもらい、アスパラギン酸になります。アスパラギン酸はグルタミン酸と入れ替わりにミトコンドリア外に運搬され、アミノ基を 2-オキソグルタル酸に渡してオキサロ酢酸に変換されます。このオキサロ酢酸は最初に NADH から電子をもらった分子です(図2の緑の経路)。
一方で、2-オキソグルタル酸はアスパラギン酸からアミノ基を受け取り、グルタミン酸になります。このグルタミン酸はアスパラギン酸と入れ替わりに輸送され、オキサロ酢酸にアミノ基を渡して 2-オキソグルタル酸に変換されます。2-オキソグルタル酸はリンゴ酸と入れ替わりに輸送されて、細胞質側へ移行します(図2緑の経路)。これで最初に戻りましたね。このようにして NADH の持つ電子はミトコンドリアマトリックス側の NAD+ に渡して NADH が産生できますので事実上 NADH が移動したことと同じことになりますね。こうして産生された NADH は電子伝達系に渡されて処理されます。
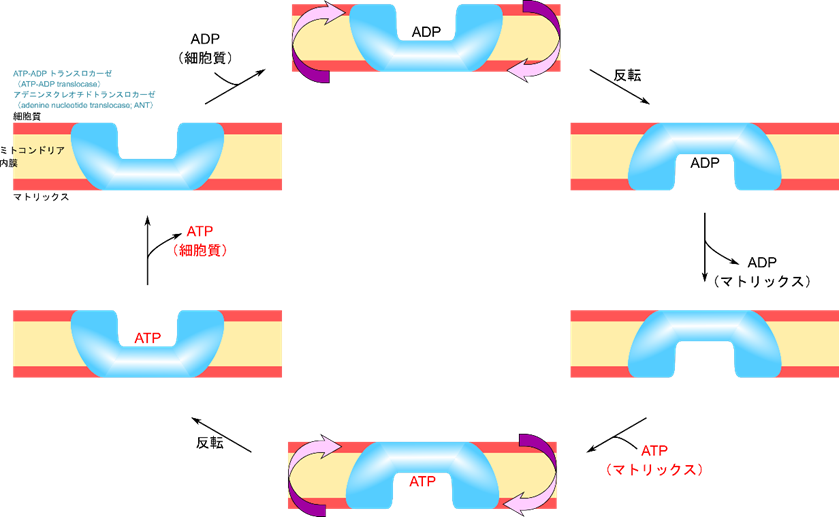
ATP-ADP トランスロカーゼによる ATP の輸送
酸化的リン酸化で産生された ATP は細胞の他の領域に運搬される必要があります。同時に ADP も ATP に再生されるためにミトコンドリアのマトリックスに運搬されなければなりません。この運搬には ATP-ADP トランスロカーゼ(ATP-ADP translocase)が関与しています。
ADP が ATP-ADP トランスロカーゼに ADP が結合すると 反転してマトリックス側に ADP が放出されます。ADP が放出された後に ATP が結合すると、再度 ATP-ADP トランスロカーゼが反転して ATP が細胞質側に放出されます。このように ADP と ATP の運搬は互いにセットになって機能します(共役しています)。ここでこの反応は
ATP3-cytosol + ADP4-matrix → ATP3-matrix + ADP4-cytosol
と書けます。電荷だけに注目すると、
細胞質の電荷 :3- → 4-(電荷の変化量:-1)
マトリックスの電荷:4- → 3-(電荷の変化量:+1)
となります。ミトコンドリアの内膜は細胞質側がプラス、マトリックス側がマイナスに荷電することで ATP 産生が行われていました。ATP と ADP の移動はこの膜電位を弱めるように作用するので、弱められた膜電位を解消するのに余分なエネルギーが必要になります。
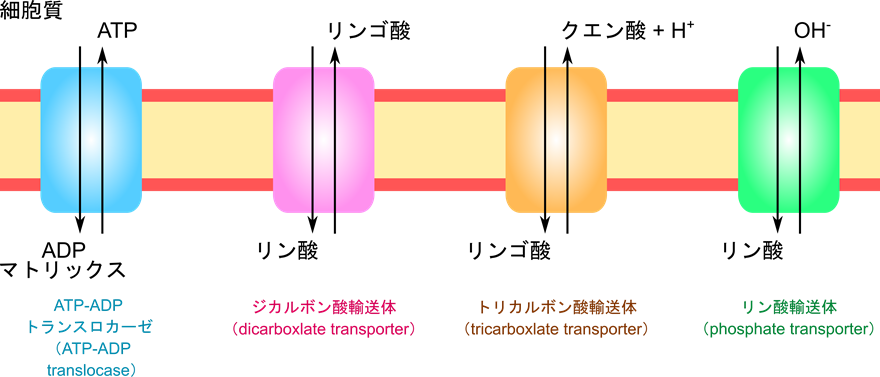
その他の物質も輸送体により輸送される
他にも ATP-ADP トランスロカーゼと似たメカニズムで様々な分子が輸送されます。以下に一部の例を記載します。
ジカルボン酸輸送体:
マトリックス→細胞質:リンゴ酸、フマル酸、コハク酸
細胞質→マトリックス:リン酸
トリカルボン酸輸送体:
マトリックス→細胞質:クエン酸とプロトン
細胞質→マトリックス:リンゴ酸
リン酸輸送体:
マトリックス→細胞質:OH–
細胞質→マトリックス:H2PO4–
※リン酸輸送体と ATP-ADP トランスロカーゼが強調して機能することで
ADP と 無機リン酸(Pi)が ATP と交換されます。
このようにミトコンドリアの内膜は基本的に物質を透過しないため、マトリックスと細胞膜間を移行させる必要のある物質は輸送体が関与してその輸送を担います。
まとめ
今回解説したように、ミトコンドリア内膜を超えて物質を移動させるためには、さまざまなシャトル分子や輸送体が関与しています。これらの仕組みは、ミトコンドリア内でのエネルギー生産や代謝において重要な役割を果たしており、私たちの体がいかに精巧に設計されているかを実感できる一例です。
参考文献
- 島原健三 (1991). 概説 生物化学. 三共出版. pp. 139-150
- Jeremy M. Berg, John L. Tymoczko, Gregory J. Gatto Jr., Lubert Stryer著、入村達郎、岡山博人、清水孝雄、中野徹訳 (2018). ストライヤー生化学 第8版. 東京化学同人. pp. 485-519
関連記事
- プロトン勾配と酸化還元反応をエネルギー的な観点から解説:生体内のエネルギー流れ
- 細胞のエネルギー工場:ミトコンドリアとATP生成メカニズム
- 電子伝達系を徹底解説:プロトン汲み出しのメカニズムとは?
- 化学浸透圧説を徹底解説:プロトン勾配とATP合成の重要なつながり
- NADHの電子はどう運ばれる?ミトコンドリア膜を越えた物質輸送を解説(←イマココ)